
")

| nota |
|
||||||||||||
|
Producción de
insulina humana
por técnicas de ADN recombinante
|
 |
||||||||||||
| Beatriz Flores | |||||||||||||
|
La diabetes mellitus es una enfermedad crónica
caracterizada por la incapacidad del individuo para metabolizar glucosa de manera normal, debido a una deficiencia total o parcial de la hormona insulina, por lo que en algunos casos, los pacientes diabéticos pueden ser controlados mediante la aplicación de insulina.
En México, toda la insulina es importada por una sola compañía productora. Se ha calculado que su consumo en 1989 fue de 56 kg y se estimó que para el año 2000 se requerirán entre 81 y 100 kg de la hormona.
Hasta 1983 toda la insulina utilizada para el tratamiento de la diabetes era extraída de páncreas de porcino y bovino, sin embargo, el uso prolongado de este tipo de insulinas provoca, en algunos individuos, una respuesta inmune. Además, la disponibilidad de páncreas de los animales mencionados es limitada, por lo que es deseable la obtención y uso de insulina humana.
La síntesis natural de la insulina comienza con la producción y procesamiento de una proteína precursora llamada proinsulina. Posteriormente, esta proteína sufre degradación en el centro, dando lugar a dos proteínas (cadenas A y B) unidas mediante dos puentes disulfuro; esta molécula es la insulina madura.
El Instituto de Biotecnología de la UNAM ha desarrollado un método de obtención de insulina humana mediante ingeniería genética de bacterias. La ingeniería genética es un conjunto de metodologías que permiten el aislamiento, caracterización y manipulación del material genético de los seres vivos. Desde 1974 ha sido posible aislar genes de algunos organismos e introducirlos en otros para su expresión. En este caso, el objetivo sería conseguir que los genes humanos, responsables de la producción de la hormona se expresaran en la bacteria. De este modo, las bacterias producirían insulina a grandes velocidades, pudiéndose escalar el modelo en tanques de fermentación para la obtención de grandes cantidades.
Existen dos rutas para la obtención de insulina humana utilizando microorganismos modificados por ingeniería genética. Una de ellas, consiste en producir por separado ambas cadenas para, posteriormente, asociarlas químicamente. La otra se basa en la producción de proinsulina que es procesada hasta insulina madura mediante métodos enzimáticos.
En el laboratorio del Dr. Francisco Bolívar Zapata se han desarrollado ambos tipos de opciones para la síntesis de la insulina humana. Este tipo de manipulación de material genético incluye varios pasos:
1) Obtención por síntesis química del gene deseado.
2) Aislamiento del gene en un vehículo de transporte tal que permita una expresión eficiente y estable en el organismo receptor.
3) Introducción del gene en el organismo receptor mediante el vehículo; en este caso se utilizó la bacteria Escherichia coli.
Este tipo de bacterias, denominadas recombinantes, pudieran ser el punto de partida para el desarrollo de procesos industriales. Sin embargo, para que esto sea posible, deberán ser cepas bacterianas que además de ser óptimas en cuanto al transporte y expresión estable del gene dentro del organismo receptor, tendrán que serlo en cuanto al procesamiento y purificación de los productos génicos expresados en el organismo recombinante.
La estrategia que el grupo del Dr. Bolívar siguió para obtener proteínas estables, fue el aislamiento de éstas unidas a otra proteína (represor C1), que se expresa normalmente en la bacteria y se caracteriza por ser de tamaño pequeño. Este tipo de proteínas se conocen como proteínas híbridas o de fusión y se acumulan en el interior de la célula bacteriana dentro de unos compartimientos llamados cuerpos de inclusión que, posteriormente, pueden ser recuperados mediante centrifugación diferencial. De ahí, las cadenas A y B o la proinsulina, son liberadas de la proteína de fusión por métodos químicos para proceder a su purificación. En este último paso, se utilizaron técnicas cromatográficas diferentes para cada una de las cadenas A y B. Una vez aisladas las proteínas A y B, se someten a reacción para que se unan químicamente, esta unión se lleva a cabo mediante proteínas presentes en ambas cadenas, que formarán los mencionados puentes disulfuro. El producto de esta reacción es la molécula de insulina activa. En el caso de la obtención de proinsulina, se libera ésta de la proteína de fusión por métodos químicos y se somete a reacción a fin de que se produzca la insulina activa. La pureza y actividad del producto se prueba en ratas.
Una vez que funciona el modelo en laboratorio, donde se manejan volúmenes de 100 mls, se prueba en lo que se conoce como planta piloto, donde es posible hacer un escalamiento a diez y cien litros, esto es, dos y tres órdenes de magnitud más, respectivamente. Este proceso requiere de una optimización en cuanto a la cantidad de biomasa que se puede recuperar.
La recuperación de las células bacterianas se lleva a cabo por filtración. Posteriormente se rompen las células por medio de una prensa francesa a fin de que se liberen los productos proteicos intracelulares. Los cuerpos de inclusión, donde se encuentra la proteína de interés, son recuperados por centrifugación diferencial. Hasta ahora, se han obtenido 1.2 grs de Insulina Activa por litro de cultivo en el escalamiento de diez litros y 0.59 grs/lt, en el escalamiento de cien litros. En ambos procesos, la eficiencia es de un 60% y la diferencia de las cantidades obtenidas entre uno y otro tanque, radica en la cantidad de biomasa obtenida: en el tanque de diez litros se han obtenido veinte grs de células por litro, mientras que en el tanque de cien lts sólo se obtuvieron nueve grs de células por litro.
El desarrollo de este tipo de tecnologías resulta muy importante para nuestro país, ya que por este medio podrán subsanarse necesidades clínicas de manera inmediata.
|
|||||||||||||
| _____________________________________________________________ | |||||||||||||
|
Beatriz Flores
Instituto de Fisiología Celular,
Universidad Nacional Autónoma de México.
|
|||||||||||||
|
_____________________________________________________________
|
|||||||||||||
|
cómo citar este artículo →
Flores, Beatriz . 1994. Producción de insulina humana por técnicas de ADN recombinante. Ciencias, núm. 34, abril-junio, pp. 61-62. [En línea].
|
|||||||||||||
 |
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Víctor M. Toledo | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Con la aparición del libro Biological Diversity of Mexico
(Ramamoorthy, et al., 1993) vino a cerrarse un primer ciclo en el estudio de la riqueza biológica del país, una etapa que se inició cinco años atrás con la aparición de mi artículo La diversidad biológica de México (Toledo, 1988), que fue la primera publicación dedicada a tratar de manera explícita este tema. En ambos casos, las publicaciones se originaron en sendos encuentros académicos realizados por iniciativa de varias instituciones tanto mexicanas, como de carácter internacional. Si esta primera etapa concluyó, fue para dar lugar a un segundo periodo. En efecto, la creación de la CONABIO (Comisión Nacional para el Conocimiento y Uso de la Biodiversidad), un organismo gubernamental, especialmente dirigido a atender este tópico, marca el inicio de lo que seguramente será la segunda etapa en el abordaje del tema. Este hecho tiene un enorme significado, porque coloca al país en una situación privilegiada, casi única, con respecto a la problemática de la conservación y uso correcto de la biodiversidad en el mundo. No solo es México uno de los pocos países en donde existe un programa de carácter nacional sobre la biodiversidad (sólo se tiene noticias de esfuerzos similares en Australia y Sudáfrica), sino que es el único “país tropical” con una reconocida megadiversidad, que cuenta con una estructura institucional de esta índole. Ello ubica al país en una situación de vanguardia, pero también obliga a quienes se encuentran comprometidos con este reto, a resolver problemas e interrogantes nunca antes visualizados, de organización académica, de teoría y metodología, de estrategia de investigación y aun de simple logística.
En efecto, la sola formulación de un proyecto nacional de investigación como el que el estudio de la biodiversidad de México plantea, supone un salto cualitativo en la investigación desde los estilos personales de los investigadores hasta los relacionados con la política científica a nivel de instituciones y del país entero. Por todo lo anterior, el presente ensayo hace una exploración de los principales retos que, para la investigación, desencadena el estudio de la biodiversidad del país, en el futuro inmediato (los noventa). Dicha exploración se realiza tomando como punto de partida un balance, o diagnóstico de los avances logrados en el campo y las nuevas interrogantes que se proyectan.
El estudio de la biodiversidad: significados epistemológico y social
El cierre del siglo encuentra a los seres humanos enfrentando una crisis ecológica de escala planetaria, uno de cuyos componentes es, precisamente, la pérdida de la “variedad de la vida” como resultado de la transformación de los hábitats, la contaminación de origen urbano-industrial (especialmente sobre los ecosistemas acuáticos continentales y marinos), la sobre explotación de los recursos y el comercio ilegal de especies. Como sucede con el resto de los principales problemas ambientales reconocidos, la pérdida de la biodiversidad ha cimbrado los círculos académicos contemporáneos, ha cuestionado buena parte de los paradigmas de las disciplinas relacionadas con el problema y ha estimulado la proposición de nuevos enfoques, métodos y conceptos. Ello ha sido así porque la conservación y uso correcto de la diversidad biótica del planeta requiere de información confiable y precisa, es decir, requiere de la investigación científica. Esta ha sido, por cierto, la manera en que, a través de la historia los nuevos campos de la ciencia se han originado: nuevas demandas sociales, esto es, la resolución de nuevas problemáticas, estimulan y crean nuevos campos del conocimiento. Desde el punto de vista epistemológico, esto se traduce, como ha argumentado Kuhn, en el cuestionamiento de una “ciencia normal”, es decir, de los paradigmas habituales. En el caso de los problemas ambientales (y la pérdida de la biodiversidad es uno de ellos), no cabe duda de que estamos viviendo un profundo reacomodo en la manera de visualizar las problemáticas y, por lo mismo, una re-configuración de los campos habituales del conocimiento científico, normalmente encargado del estudio del tema.
Desde otra perspectiva, puede afirmarse que el estudio de la biodiversidad está dando lugar a un nuevo campo o enfoque del conocimiento (¿la biodiversología?) o a lo que Soulé ha llamado (al referirse a la biología de la conservación) una “disciplina de crisis” (crisis discipline), y en la cual ante la urgencia de datos, “…one must act before knowing all the facts; crisis disciplines are thus a mixture of science and art, and their pursuit requires intuition as well as information”. Cualquiera que sea el destino de esta nueva forma de abordar el problema, queda claro que la necesidad de salvaguardar el patrimonio biótico del planeta y sus espacios (países, regiones, localidades) está replanteando el significado normal de las ramas de la biología, tradicionalmente encargadas de atender estos asuntos. En efecto, lo que presenciamos hoy en día es una reformulación del fenómeno evolutivo (esta vez por sus implicaciones prácticas) y de las tres ramas que se ocupan de interpretarlo (la taxonomía, la ecología y la biogeografía). Esta reconfiguración surge como una respuesta a problemas y preocupaciones concretas del mundo contemporáneo, tales como la pérdida de genes y organismos, el uso y manipulación de genes y especies con utilidad real o potencial, y el mismo equilibrio ecológico del sistema planetario. Estas disciplinas se encontraban en una especie de “encantamiento academicista”, el cual tendía a mantenerlas confinadas al aislamiento, es decir, como estancos separados. Pero el nuevo concepto de biodiversidad ha tenido la virtud de sacarlas de ese estado para reunirlas y ponerlas en acción, de manera integradora, en torno a un problema concreto. Es decir, las ha vuelto un conocimiento, socialmente demandado, que se requiere con urgencia. Parece entonces que estamos ante el advenimiento de un área del conocimiento con una estructura sintética, ecléctica, pragmática y multidisciplinaria. Veamos cómo toma cuerpo esta situación, en el caso del estudio de la biodiversidad de México, no sin antes hacer una rápida revisión de lo que este concepto implica.
El concepto de biodiversidad: definiciones, explicaciones y mediciones
El empleo del término diversidad biológica es reciente; surgió indisolublemente ligado a las instituciones académicas y organismos nacionales e internacionales dedicados a la conservación biológica, y como un concepto sintético que incluye por igual enfoques de la taxonomía, la ecología y la biogeografía. Es al mismo tiempo un concepto que implica una finalidad práctica: la evaluación de los ambientes naturales (y sobre todo) perturbados del planeta. En tal sentido, el concepto de biodiversidad de carácter tridisciplinario y pragmático resulta diferente del concepto de diversidad de especies, postulado hace casi cuatro décadas como un rasgo estructural ecosistémico, a partir de la aplicación a la ecología, de la Teoría de la Información (por ejemplo, Margalef, 1957 y 1968). En varios sentidos, el término de biodiversidad conlleva un significado amplio, de tal forma que, como bien señala Pielou (1991), tal concepto abarca varios diferentes tipos de diversidad: genética, específica, estructural, ambiental y ecosistémica (véase también Lévèque & Glachant, 1992 y di Castri & Younés, 1990).
Para complicar el cuadro, el concepto de biodiversidad, que implica la medición de la riqueza biótica en un espacio y un tiempo determinados, dado su significado práctico, también conlleva un componente geopolítico. Por ello, el empleo del término megadiversidad biológica, introducido por Mittermeir, así como su utilización como un elemento más de la negociación internacional (en la Cumbre de Río), otorga a este concepto una dimensión cualitativamente nueva, que lo deja muy lejos del concepto de diversidad empleado, de manera “doméstica”, por los investigadores de la ecología (biológica). Usher de alguna forma coincide con esta apreciación cuando afirma: “Biodiversity is therefore an integration of natural history (or knowing what species there are), measurement (or how to asses what we mean by diversity and how to compare sites) and use (or the realization that many species are important for the long term survival of Homo sapiens)”. Por todo ello, el concepto de biodiversidad ha sido ya integrado al vocabulario normal del pensamiento conservacionista (por ejemplo Wilson, 1988; McNeely, et al.,1990).
El carácter cualitativamente nuevo del concepto de biodiversidad (por sobre el de diversidad de especies) se vuelve determinante cuando se revisan tanto las diferentes elaboraciones teóricas que tratan de explicar el fenómeno, como las fórmulas propuestas para medirlo. Que los patrones de biodiversidad rebasan el ámbito meramente ecológico, ha sido puesto en evidencia por la investigación empírica de las últimas décadas (véase Brown, 1988), que muestra la insuficiencia de los modelos basados en principios tales como la disponibilidad de recursos, predación, competencia, equilibrio entre especiación/inmigración y extinción/emigración y estructura o productividad del hábitat, formulados desde la teoría ecológica (Mac Arthur, 1972; Terborg, 1974). Otros factores, como aquellos utilizados por la biogeografía para explicar patrones o tendencias en la distribución espacial de las especies (por ejemplo, procesos históricos, barreras geográficas, refugios, etcétera), resultan de mayor importancia sobre una cierta escala geográfica. Más fértil ha sido por lo tanto la explicación de fenómenos particulares, basados en evidencias empíricas, que los intentos por crear una teoría general. La explicación de los patrones de biodiversidad encontrados, varía, por lo menos, en función de la escala espacial, el tipo de organismo estudiado, la condición del hábitat (terrestre versus acuático o marino), etcétera. Muy notable es la importancia de la escala espacial del estudio, un hecho que hoy en día obliga a repensar los dominios de las disciplinas y aun sus definiciones.
En el caso de la medición de la biodiversidad sucede algo semejante. Si bien se han realizado propuestas sofisticadas para medir la diversidad de especies (véanse los libros de Pielou, 1975 y de Magurran, 1988 y la última contribución sobre el tema de Margalef, 1991), después de una cierta escala, la fórmula más utilizada sigue siendo la más simple: el cálculo de la riqueza (número de especies) por unidad de superficie. Esto es así porque en la práctica los cálculos más complicados (por ejemplo, los índices de Shanon-Weaver o Simpson) sólo son factibles sobre escalas muy finas, con las que puede obtenerse la información requerida (véase una discusión sobre este tema en Brown, 1988). Por otro lado, algunos autores se preguntan si una simple cifra logra expresar cabalmente el fenómeno, y si no resulta conveniente buscar otras formas para representarlo. Por ejemplo, siguiendo lo sugerido por Margalef (1968), es posible elaborar espectros de diversidad, los cuales expresan la tendencia del incremento de este parámetro, conforme aumenta el tamaño de la muestra o de la colección (figura 1). De enorme interés es la reciente propuesta de Pielou (1991) que consiste en buscar, por una vía diferente un índice de biodiversidad de carácter totalizador (esto es que incluya todos los grupos de organismos presentes en un espacio determinado). Pielou (op. cit.) sugiere utilizar un vector de diversidad (diversity vector) que para ser útil, debe cumplir con dos propiedades: tendrá que ser elaborado a partir de información de campo de fácil obtención y, también que tendrá que ser rápidamente comprendido por los no especialistas. El mismo autor se inclina por el uso de diagramas de diversidad, en los que deberían quedar expresados ciertos patrones de la biodiversidad de un ecosistema o comunidad, tales como el espectro de formas de vida vegetales y otros fenómenos.
Más allá de las diferentes interpretaciones de la biodiversidad y de sus formas de medirla, nos encontramos con tres momentos o etapas del proceso cognoscitivo. El estudio de la biodiversidad, como el de cualquier fenómeno de la realidad natural o social, pasa necesariamente por tres fases bien definidas: una descriptiva, una interpretativa (o analítica) y una última predictiva. En todos los casos se trata, en última instancia, de descubrir y describir patrones, es decir, regularidades en los fenómenos que se abordan. Los apartados que siguen hacen una exploración acerca de algunos de los retos que esperan a los estudiosos de la biodiversidad en México. Aunque he tomado como universo exploratorio el caso particular de un país, las aportaciones y propuestas tienen un significado general; es decir, deberían ser válidos para otros países que buscan igualmente la comprensión de este fenómeno, en una escala nacional.
El primer paso necesario: los inventarios biológicos
La búsqueda de patrones de biodiversidad es imposible si no se cuenta con una estructura básica de conocimiento taxonómico. Esto es aún más cierto cuando se trata de buscarlos a una escala nacional. Ello implica no solamente el disponer de un conocimiento confiable acerca de las especies que habitan un espacio determinado (en este caso México), sino de sus distribuciones a través de ese espacio. La ausencia de este conocimiento hace aparecer como patrones aparentes lo que no son sino artificios de colecta (por ejemplo, especies que aparecen como endémicas cuando en realidad no existen colectas suficientes). El problema reside entonces en saber en qué momento un inventario (que no es sino una muestra de escala nacional) alcanza el mínimo de confiabilidad. Esto depende, por supuesto, del grupo de organismos que se estudia y del espacio geográfico (en este caso un territorio nacional). Examinemos el caso de las plantas mexicanas, ya que ello nos dará oportunidad de atender el grado de dificultad que esto encierra.
En México no conocemos aún el total de especies de la flora fanerogámica, la cual se estima que aloja entre un mínimo de 23000 especies y un máximo de 30000 (Toledo & Rzedowski, 1993; Toledo, 1993b). Como ningún otro país en Latinoamérica, la colecta de plantas se incrementó en las últimas décadas al pasar de 566000 especímenes en 1974 (alojados en 18 herbarios nacionales) a 2100000 en 1990 (en 71 herbarios) (Toledo & Sosa, 1993). En un trabajo reciente, Campbell postuló, de manera arbitraria, que un índice de densidad de colecta mínima para los países tropicales, debería de ser de 100 especímenes por 100 km2. Hacia 1990, México presentaba un índice de 107. Si a las colecciones nacionales se agrega el número de especímenes depositados en herbarios extranjeros, que se calcula es de 1900000 (Toledo & Sosa, 1993), el país quedaría con un gran total de unos cuatro millones de especímenes colectados. Esto deja al país con un índice global de densidad de colecta botánica más que suficiente: sobre 200 especímenes por cada 100 km2. Sin embargo la realidad es otra. A pesar de que existen 16 floras regionales terminadas, o en proceso, que abarcan el 70% del territorio nacional, y del que ya está descrito el 75% de las especies consignadas en listados florísticos —checklists— (Sosa & Dávila, 1993), el país no cuenta aún con un listado de especies a escala nacional; es decir, no se conoce aún su diversidad florística. Todavía más, algunas de las que se suponen son ya floras terminadas, han sido enriquecidas recientemente por nuevos esfuerzos de colecta. Este es el caso de la flora de Baja California, a la cual Delgadillo J. R. agrega 800 especies; la de Quintana Roo, a la cual Villanueva y Cabrera adicionan 175 especies, y la de Chiapas a la cual Esteban Martínez (comunicación personal) le agrega ¡1000 especies! Ello resalta la insuficiencia que existe en las colectas y que prevalece aún en los territorios o regiones. Por otra parte, revela el carácter limitado de los índices de densidad que suponen que las colectas han sido realizadas de manera uniforme en el espacio. En análisis recientes sobre la cartografía de las colectas, se puede ver que muchas veces los colectores se concentran en los sitios más accesibles o con menor grado de dificultad (una muestra de esto es el conocido sesgo de la colectas junto a las carreteras).
El ejemplo anterior muestra la necesidad de evaluar con precisión el estado que guardan los inventarios biológicos, lo que nos obliga a crear índices o fórmulas para evaluar el grado de avance de un inventario biológico determinado y esto, a su vez, hace necesario el usar modelos teóricos que evalúen la relación que se establece entre el “esfuerzo del conocimiento” y el universo por conocer. Todo esto nos hace regresar al problema de la escala, pues lo que aquí denominamos inventario, no es sino una muestra a escala nacional. Entonces, ¿cómo formular modelos que sean igualmente válidos para unas cuantas hectáreas (que es la escala a la que trabajan los ecólogos) y para espacios microregionales, regionales, estatales y de todo el territorio nacional? El punto de partida es, como en otros muchos casos, los avances logrados desde la teoría ecológica (véase Soberón & Llorente, 1993), que hoy son la porción más sólida del terreno. Sin embargo, se necesita mucha intuición y arte (invocando a Soulé), para matizar estas formulaciones, para poder utilizarlas en cada una de las diferentes escalas. Por ejemplo, la espacialización del “esfuerzo de colecta”, que en un modelo esencialmente ecológico se expresa por la relación que existe entre el número de nuevas especies encontradas con el tiempo empleado por un número determinado de investigadores (Soberón & Llorente, op. cit.), puede quedar expresado, en otra dimensión, a través de mapas. En otros casos, la simple revisión histórica de la bibliografía, o de las colecciones, han ofrecido una idea del estado actual del conocimiento. Este ha sido el caso de los mamíferos (Ramírez-Pulido & Britton, 1981; Ramírez-Pulido & Müdespacher, 1987), los musgos (Delgadillo, 1990), las compuestas (Cabrera-Rodríguez & Villaseñor, 1987) o los sifonáptera (Morales & Llorente, 1986). Lo anterior llevaría a reconocer la necesidad de crear una cartografía de esfuerzo de colecta para cada grupo de organismos, sobre diferentes escalas espaciales, y elaborado bajo normas, criterios y estándares bien establecidos. Para ello tendrían que crearse comités de especialistas para cada grupo de organismos, que serían los responsables de realizar esta tarea. La teorización deberá ser un juego de proyecciones espaciales, a partir de áreas bien conocidas, combinado con técnicas de distribución geográfica de las especies, etcétera. La tarea puede ser relativamente fácil en grupos bien colectados y de relativamente pocas especies, como lo son los vertebrados (especialmente los mamíferos) y quizás las plantas, pero esto tiende a complicarse en grupos tales como los insectos. Tendrán que buscarse otros métodos a partir de las colecciones mismas y sus registros; es decir evaluando el esfuerzo del conocimiento mediante formas de cuantificación de especímenes en herbarios y museos. De cualquier forma se hace necesario inducir y formalizar las investigaciones teórico-metodológicas en torno al grado de avance de los inventarios biológicos, en todas sus facetas y dimensiones. Ello debería permitir la uniformidad metodológica con la intención de tener una panorámica, bien sustentada, del grado de conocimiento básico sobre las especies de México y las necesidades de investigación en el futuro.
La segunda lección que se desprende del examen realizado con las plantas, es que la determinación de la riqueza biológica del país, en lo que concierne a su primera etapa (los inventarios), debe ser obligatoriamente un esfuerzo no solo meta personal sino multiinstitucional. La realización y terminación de los inventarios biológicos, es entonces la primera prioridad en todo proyecto que intenta conocer la biodiversidad de un país. El caso de las plantas resulta de nuevo ilustrativo. Todo indica que hay que dar el “paso final” para integrar no solo un listado florístico del país, sino también un banco de información que incluya registros de colecta (etiquetas), distribución geográfica, caracteres principales, usos, imágenes de la especie, etcétera. (Sosa & Dávila, 1993). La simple acumulación del número de especies derivados de las floras regionales, los endemismos, los nuevos registros, etcétera, sacados de la literatura nos ofrecen ya una primera cifra de lo hasta ahora conocido: más de 27000 especies (Cuadro 1). Esta etapa de consolidación de la flora de México requiere sin embargo del consenso y la intención de las múltiples instituciones involucradas en el asunto y de sus cuerpos de investigadores; una iniciativa que debería ser impulsada por el Programa Flora de México, con más de una década de existencia.
Patrones taxonómicos
Un primer conjunto de patrones pueden detectarse de manera aespacial, es decir, circunscritos a la simple numerología de las especies por grupos de organismos y a sus relaciones entre ellos. A éstos podríamos denominarles patrones taxonómicos. El primer patrón es el número de especies que existen en México, en relación con el número total descrito para el mundo. Por lo que indican los grupos de organismos mejor conocidos, México parece contener entre un 10 y un 12% del total de la biota del mundo. Este porcentaje se ajusta al caso de las plantas con flores (y entre ellas la familia Leguminosae y las plantas acuáticas), los musgos, los mamíferos, las aves, los reptiles, las abejas y las mariposas. Las dos excepciones que saltan a la vista son el caso de los anfibios y el de los peces marinos, con solo un 7%. Para el caso de las algas marinas, puede esperarse que también se ajuste el patrón, ya que hasta ahora existe un primer listado de 2000 especies (básicamente macroalgas), que si aumenta en un 50% cuando se le agreguen las microalgas bentónicas y el fitoplancton y otros grupos no estudiados (Novelo, comunicación personal), darían un total de hasta 3000 especies. Esta cifra representaría algo más del 10% del total de especies de algas marinas reportadas en el mundo (26900; aunque otros autores dan 40000). De ser correcto este porcentaje, el reto que le espera a los biólogos mexicanos es el de documentar entre 300000 y 3 millones de especies de organismos (dependiendo de la posición que se tome en la actual polémica sobre el total de organismos que se considera pueden existir en el mundo; véase: Wilson, 1988; Stork & Gaston, 1990; May, 1992). Del resto de los grupos poco puede decirse, ya sea porque los inventarios aún están en proceso, o porque existe demasiada controversia entre los taxónomos, acerca de la categorización de las especies.
Como sucede a nivel mundial, destaca la necesidad de enfocar el esfuerzo de los inventarios, hacia el área de los insectos e invertebrados marinos, ya que juntos representan ¡más del 90% de la diversidad biológica del planeta! En el caso de los insectos podrían aventurarse algunas estimaciones de importancia en la estrategia de estudio, mediante su relación con las especies de plantas (familias y géneros) o de animales (por ejemplo, el caso de las pulgas, en relación a los mamíferos; véase Morales & Llorente, 1986). Esto permitiría medir la dimensión del esfuerzo que se requiere para completar el inventario de ciertos taxa. Los grupos de microorganismos (hongos, bacterias, virus y protozoarios) son de enorme importancia debido al papel clave que juegan en los procesos del metabolismo ecosistémico (captación de energía, fijación de nitrógeno, descomposición, simbiosis, calidad de los suelos, detoxificación, etcétera) ya que son, a nivel mundial, los organismos con los inventarios menos desarrollados. En México se debería poner mayor énfasis en su estudio y catalogación, por ser de carácter estratégico en los sistemas de producción de alimentos y medicinas, así como para la biotecnología. I. La dimensión geográfica: Patrones espaciales
Un primer abordaje de la biodiversidad, en relación con el espacio, se logra mediante la simple delimitación del área de las especies, es decir, de su distribución geográfica (la cual es cartografiable). La aerografía, que es el estudio de las estrategias geográficas de las especies (Rapaport, 1975), es la expresión más acabada del estudio teórico de la diversidad biológica en un espacio ecológicamente abstracto. En este caso, la búsqueda de patrones se logra mediante la correlación, a través de las técnicas de regresión estadística, entre el número de especies y las variables del medio físico. De esta forma se obtienen gradientes de la diversidad, como resultado de la variación de uno o más factores físicos. Los patrones más conocidos son, entre otros, los relacionados con los gradientes latitudinal, altitudinal, de aridez, de salinidad y de profundidad (en el medio marino), de concentración de sustancias en el suelo, aire y agua, etcétera (véase Brown, 1988). Como sucede con la mayor parte de los análisis efectuados sobre la biodiversidad, estos patrones han sido descubiertos en grupos de macroorganismos terrestres, pero han sido muy poco explorados en el medio acuático (especialmente el marino) y entre grupos de pequeña talla (por ejemplo insectos, hongos o musgos).
En México son varios autores los que han encontrado patrones geográficos de la diversidad, durante el estudio de algunos grupos de organismos. Estos patrones han sido relacionados con la latitud, la elevación, las provincias bióticas y biogeográficas, las regiones fisiográficas y naturales, ciertos sitios geográficos y, por supuesto, las unidades geopolíticas representadas por los estados de la república (véase Rzedowski, 1993; Llorente & Martínez, 1993; Ayala, et al., 1993; Flores-Villela, 1993; Escalante, et al., 1993; Fa & Morales, 1993; Riba, 1993; Toledo, 1982).
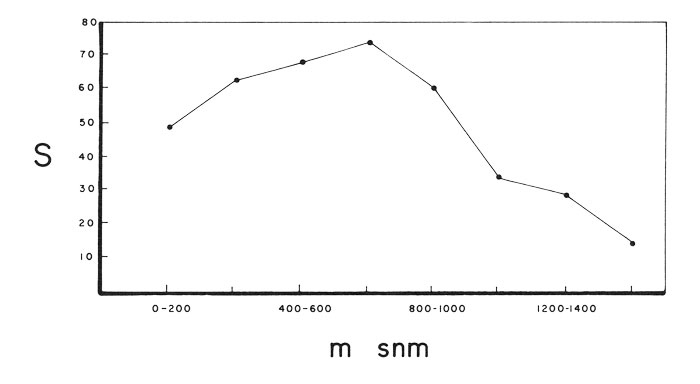
Una técnica que frecuentemente se ha utilizado, es el simple conteo de las especies, encontradas en el territorio dividido en cuadrantes de diferentes tamaños. Para México, es de enorme interés el estudio comparativo de los patrones geográficos entre grupos diferentes de organismos. Por ejemplo, Fa & Morales (1993:351) encuentran similitudes entre las provincias bióticas que resultan del análisis de aves, mamíferos, reptiles y anfibios. Y en la perspectiva geopolítica las evidencias provenientes de diferentes grupos indican que Oaxaca, Chiapas y Veracruz son los estados más ricos de la República (figura 2). Sin embargo, el estudio de las anomalías o singularidades en la distribución geográfica de las especies un aspecto, es el que se antoja fascinante. En efecto, la ubicación latitudinal de la República mexicana, su compleja fisiografía y climatología y su historia particular, hacen de México un escenario sui generis. Por ejemplo, el patrón latitudinal que establece un aumento progresivo de especies hacia el ecuador y que parece ser de carácter universal entre los macroorganismos (Brown, 1988), no siempre se ajusta al caso de México, ya que se presenta una muy desusada riqueza de especies entre los 19 y 20° de latitud norte, debido al del Eje Neovolcánico Transversal. Este fenómeno ha sido señalado por diversos autores (Halffter, 1964; Rzedowski, 1978; J. R. Delgadillo, 1987 y especialmente Fa, 1989) en el estudio de grupos, tales como los insectos, las plantas, los musgos y los mamíferos. De manera similar, el patrón de disminución de especies, conforme aumenta la altitud, se ve modificado por la enorme riqueza del bosque mesófilo de montaña, una comunidad vegetal que se distribuye entre los 900 y 2000 msnm y que es el sitio de encuentro entre las especies neárticas y neo tropicales. Este fenómeno, que fue sugerido desde principios del siglo por Gadow (1907-1909), parece operar no solo entre las especies de plantas (figura 3), sino entre algunos grupos animales, como las aves (figura 4). Finalmente, el incremento de especies de plantas, conforme aumenta la humedad, que ha sido un patrón postulado por Gentry (1988 y 1992) basándose en estudios realizados en las zonas tropicales de los tres continentes, parece ser un patrón que no opera en México, ni en la escala de grandes áreas (Rzedowski, 1993), ni sobre la comparación de sitios específicos (Toledo, 1982). En ambos casos parecen tener mayor importancia los sucesos de tipo histórico.
II. La dimensión ecológica: Patrones espaciales
De enorme importancia estratégica es el distinguir entre los patrones de la biodiversidad en el espacio, tomado de manera ecológicamente abstracta (apartado anterior), y el espacio conceptualizado como un conjunto de unidades ecológicas discretas (las comunidades de organismos y su entorno abiótico). Podría decirse que en este último, el observador enfoca su objeto para distinguir con precisión el conjunto de especies representado por la comunidad biótica y su entorno no vivo, que media entre los organismos estudiados y los factores del medio físico. Ello nos lleva a reconocer el concepto de hábitat, y a todo espacio (terrestre o acuático), como un conjunto de hábitats. Por lo tanto, los patrones de diversidad se exploran, ya no solo en función del área de la especie (es decir, su distribución geográfica), sino de su presencia o ausencia en los hábitats distinguidos en el espacio. En sentido estricto, los criterios que se requieren para definir un hábitat, se establecen basándose en la naturaleza específica de los organismos que están bajo estudio. Las características y dimensión del hábitat son diferentes para, por ejemplo, los vertebrados o las especies de árboles, que para los insectos, las algas o los moluscos. Por ello habría que formular un sistema jerárquico de hábitats en función de la escala. Sin embargo, parece lógico pensar que, en una primera instancia, el espacio puede ser dividido en grandes macro (¿mega?) hábitats y que los (meso y micro) hábitats incluidos en ellos representan subconjuntos de los primeros; es decir que son variaciones dentro de intervalos mayores.
Los hábitats terrestres también son definidos de manera diferente a los acuáticos, y dentro de estos últimos habría que distinguir entre los de las aguas continentales y los marinos. Por ejemplo, mientras que en la porción terrestre, para la casi totalidad de los organismos, el espacio es de carácter bidimensional, en el medio acuático los organismos nadadores (nekton) se mueven en tres dimensiones.
De acuerdo con lo anterior, la búsqueda de patrones ecológicos de la biodiversidad en el espacio requiere de la delimitación de los hábitats terrestres y acuáticos. En efecto, sólo mediante el manejo común de una tipología de hábitats será posible explorar patrones generales del comportamiento, de la biodiversidad de diferentes grupos. En el caso de la porción terrestre, existe una franca tendencia a reconocer en el clima y la vegetación, los dos criterios centrales para poder definir los grandes hábitats. De esta forma, varios autores han dividido el territorio nacional basándose en estos criterios, durante el estudio de la cultura (West, 1964), la flora (Rzedowski, 1978), la fauna de vertebrados (Flores & Gerez, 1989), los “ecosistemas” (Pérez-Gil, et al., 1984), la producción de alimentos (Toledo, et al., 1985) y la producción rural (Toledo, et al., 1989). Siguiendo esta corriente, Toledo & Ordoñez (1993) han propuesto una división del territorio en 7 hábitats principales (o zonas ecológicas), a través de la cual es posible reconocer grandes unidades ambientales en el espacio. Esta tipología se basa en el agrupamiento de varios tipos de vegetación bajo criterios fisionómicos y biogeográficos y en sus relaciones con los principales tipos de clima (véase García, 1989), y coincide en lo general, con la realizada por otros autores (Rzedowski, 1993; Flores, 1993). Por sus dimensiones, estos hábitats terrestres resultan equivalentes a los conceptos de región natural o bioma, utilizados por otros autores.
La figura 5 muestra la manera como estos hábitats son definidos y cómo se distribuyen a través de la República mexicana. Veamos ahora cómo se comporta la biodiversidad en estos grandes hábitats analizando dos grupos bien conocidos de organismos: la flora fanerogámica (siguiendo las estimaciones de Rzedowski, 1993) y las aves (de acuerdo con Escalante et al., 1993). En el caso de la flora, la mayor riqueza de especies se encuentra en el hábitat templado subhúmedo (zona 4), esto es, con bosques de pino-encino, seguido del hábitat árido y semiárido (zona 5) y el trópico subhúmedo (zona 2). Si evaluarnos la riqueza relativa (número de especies por unidad de superficie), el hábitat templado húmedo (zona 3) queda en primer sitio: 3000 especies de plantas en solo el 1% del territorio del país. La misma situación se presenta en el caso de la riqueza en endemismos, con la salvedad de que el hábitat árido y semiárido se coloca en segundo sitio (con unas 3600 especies endémicas). Esta primera aproximación revela patrones interesantes, entre los que destaca la menor riqueza global (es decir, en su diversidad delta, véase la sección siguiente) del trópico húmedo, un hábitat que ha sido privilegiado a nivel mundial, en lo que respecta a su diversidad alfa —o de sitios específicos— (véase un reclamo similar en Redford, et al., 1990). Este fenómeno requiere de una explicación convincente, la cual se encuentra ligada a la situación de insularidad, contigüidad, tamaño y forma de las áreas que conforman cada hábitat dentro del territorio mexicano. Para el caso de las aves (figura 6), existen patrones diferentes a los encontrados en las plantas. Por ejemplo, la mayor riqueza de especies se localiza en el hábitat tropical húmedo, seguido muy de cerca por el hábitat (4) templado subhúmedo y por el hábitat (3) tropical subhúmedo. El tercer rango lo comparten la zona (5) árida y semiárida y el hábitat (3) templado húmedo. Sin embargo, en el porcentaje de endemismos, la avifauna de México se comporta de manera muy similar al caso de la flora. Las especies endémicas se encuentran en la zona árida y semiárida, seguido de los hábitats templado subhúmedo y tropical subhúmedo. De nuevo, como sucede con las plantas, en el trópico húmedo se presenta un bajo porcentaje de endemismos. Se deberían realizar comparaciones de este tipo, una vez que se cuente con la información de otros grupos, lo que permitirá detectar los factores que determinan la biodiversidad del país sobre una escala amplia.
La dimensión ecológica de la biodiversidad no se restringe, por supuesto, a la sola búsqueda de patrones en relación con los hábitats, sino que existe también una perspectiva que está muy relacionada con el papel que juegan los conjuntos de organismos en el entramado ecosistémico (Di Castri & Younes, 1990). Por ejemplo, ¿cuál es el papel funcional de la biodiversidad en relación con la viabilidad y reproducción de los ecosistemas terrestres y acuáticos? La relación de la biodiversidad con el funcionamiento ecosistémico la determina el tipo de organismos (con un énfasis en el papel de los microorganismos); la función de ciertas especies (keystone species) dominantes o claves, o grupos de éstas; el nivel de redundancia funcional de las especies; su papel en las cadenas tróficas, etcétera. El estudio de estos aspectos no es por supuesto una tarea nada fácil, ya que depende del grado de conocimiento fino que se tenga de la ecología de poblaciones de las especies y del conjunto biótico. Por ello resulta conveniente la propuesta de Field y Vázquez-Yanes, de estudiar “sistemas modelos”, en vez de intentar un análisis completo de todas las especies que contribuyen a la diversidad funcional del ecosistema. Para ello, dichos autores ofrecen un ejemplo en el análisis de las especies de Piper, en una comunidad tropical húmeda de México (Los Tuxtlas, Veracruz). Finalmente, la biodiversidad se encuentra ligada, de alguna forma, a la heterogeneidad o variedad ambiental o paisajística, y este aspecto constituye otra dimensión que debe estudiarse en el futuro.
III. El efecto de la escala: Patrones espaciales
Como es de esperarse, los valores de la diversidad biológica están fuertemente relacionados con la escala espacial en la que se realiza la muestra. La diversidad se incrementa, por supuesto, en la medida que se aumenta el tamaño de la muestra. Sin embargo, surge una interrogante crucial que es: cómo varía la tasa de incremento de especies en las diferentes escalas del espacio (tanto terrestre como acuático), y si existen patrones de escala en función del grupo de organismos estudiados. Hasta la fecha, la única distinción espacial de la diversidad de las especies, que se ha empleado y que tiende a reproducirse como un dogma en todos los estudios sobre el tema, es la propuesta por Mac Arthur hace casi tres décadas, que más tarde desarrolló Whittaker. La diferenciación espacial que allí se propone diferencia entre la diversidad alfa (o dentro de un hábitat) en áreas que por lo general no sobrepasan la hectárea; la diversidad beta (o entre hábitats), que es el resultado de la comparación de la diversidad alfa encontrada en diferentes situaciones; la diversidad gama, que se refiere a la encontrada en regiones o paisajes; y la diversidad delta que registra la heterogeneidad regional.
Shmida y Wilson (1985), han hecho notar que las tres primeras dimensiones de la diversidad coinciden con los enfoques que respectivamente hacen la ecología (o biología) de poblaciones, la fitosociología y la biogeografía clásica. Nuevamente todo esto queda dentro de una dimensión espacial, exclusivamente terrestre (lo cual deja a los medios acuáticos como áreas casi vírgenes para probar este tipo de enfoques). El reto actual, cuando los investigadores cuentan con nuevas tecnologías (como la que encierran los Sistemas de Información Geográfica) y más información sobre la distribución espacial de las especies, es el de poner a prueba esta categorización espacial que es la que se ha venido utilizando. Una contribución importante para la comprensión teórica de los diferentes factores que inciden a cada escala espacial, es la de Schmida y Wilson. En México, esta exploración permitirá precisar las aportaciones que hace cada escala espacial a su biodiversidad total.
Más allá de la fascinación teórica que encierra, los efectos de la escala espacial sobre la riqueza de especies, adquiere una importancia práctica, en términos de la definición de las áreas requeridas para conservar conjuntos de especies, aspecto que está siendo explorado con detalle en la región amazónica (Lovejoy, et al., 1986), y que a su vez, permitirá diseñar con más confiablidad algunas estrategias de conservación, basadas en las redes de reservas de diferentes tamaños y con distintas ubicaciones, lo que en sí es un aspecto crucial, ya sea en una escala nacional o en la megadiversidad. Dentro de esta perspectiva, los conocimientos acumulados en México sobre algunos grupos de organismos (especialmente vertebrados terrestres y plantas) resultan de enorme interés para comenzar a evaluar estos posibles patrones. Aquí de nuevo la complejidad paisajística del país opera como un laboratorio natural, casi único. Tomemos como ejemplo la diversidad de la flora del trópico húmedo (es decir, las especies de plantas de las selvas altas-medianas perennifolias y subperennifolias, o lo que Rzedowski llama el bosque tropical perennifolio). De acuerdo con Rzedowski, la riqueza estimada de esta flora en el país es de 5000 especies. Un banco de información elaborado durante los últimos años por el autor y por sus colaboradores, registra un total de 3100 especies, cifra que se obtiene al acumular las especies registradas en cinco regiones estratégicas del país. A nivel regional (es decir en áreas que oscilan entre 500 y algunos miles de hectáreas), la riqueza de especies varía (dependiendo de la ubicación geográfica), entre 800 y 1600 (cuadro 2). Por otro lado, de las 3100 especies, un poco más de 2000 son “exclusivas” y sólo 1000 son comunes a dos o más sitios. Lo anterior significa que, con la preservación de 5 áreas regionales de unas mil hectáreas cada una, ¡se estaría protegiendo el 60% del total estimado de la flora tropical húmeda del país! La ausencia de datos a escalas más finas impide que se puedan realizar otras apreciaciones, aunque en una sola hectárea se encuentran 267 especies (sin incluir epífitas) y en 1000 m2 más de 100 especies. La misma exploración, esta vez con los árboles (Cuadro 2) revela que una tercera parte del total de especies arbóreas registradas (690) pueden hallarse a escala regional y una sexta parte en muestras de ¡solo una hectárea! Finalmente 2000 m2 parecen contener la mitad de las especies de árboles que se encuentran en una hectárea. Los resultados de los estudios que se realizaron entre otros grupos de organismos del Trópico húmedo, también sugieren algunos patrones. Por ejemplo, bastan unas 500 hectáreas para tener la representación de la mayor parte de la diversidad de aves de una selva tropical (véase el caso de Los Tuxtlas, con 315 especies en Coates-Estrada y Estrada, 1985), y un solo sitio de la Lacandona (Chajul), contiene el 90% de las mariposas del trópico húmedo de México (543 especies, véase De la Maza y De la Maza, 1985). Estos posibles patrones, por simples que parezcan, adquieren la jerarquía de criterios cuando se van a tomar decisiones que conciernen a la conservación y uso de la biodiversidad.
Patrones de la destrucción: factores que afectan los escenarios de la biodiversidad
Hasta ahora hemos examinado los posibles patrones de la biodiversidad, bajo el supuesto de que estos existen en condiciones donde la perturbación humana es nula. Por desgracia, esta situación se está volviendo cada vez más hipotética. Hasta la fecha, el sistema de parques nacionales, y otras áreas protegidas, se extiende por sólo algo más del 3% de la superficie terrestre del planeta, mientras que se estima que el 95% de la porción terrestre está ocupada por áreas agropecuarias, forestales y para asentamientos humanos (Pimentel et al., 1992). Por lo tanto, estamos obligados a encarar el conjunto de factores extra-biológicos que determinan la existencia de los organismos, los cuales se encuentran bajo el asedio de las sociedades humanas y viviendo una suerte de “juego existencial”. La afectación (gradual o catastrófica) de los hábitats naturales y de otras formas de extinción de especies es, pues, un tema que también debe ser analizado. Ello nos obliga a ubicar y entender los patrones de la destrucción, es decir, el conjunto de factores sociales, económicos, demográficos, culturales, etcétera, que provocan la desaparición individual o masiva de las especies. Dado que la extinción o desaparición de las especies vegetales y animales tiene como causa principal la destrucción de los hábitats naturales; toda política dirigida a mantener la biodiversidad requiere de información precisa y confiable acerca de los procesos de transformación de los hábitats; sus causas, características, tendencias y proyecciones. Por lo común, la transformación de los hábitats naturales se provoca por el establecimiento de diversos fenómenos de carácter social, cultural y económico, tales como el incremento y la expansión de los núcleos humanos, los diferentes modos de percibir y utilizar los recursos naturales, o la expansión de la producción rural (agropecuaria, forestal, pesquera, extractiva) y sus efectos (por ejemplo, incendios forestales, contaminación por agroquímicos, fragmentación de masas de vegetación, etcétera).
En el caso de México, país en el que existe una fuerte presión sobre la flora y la fauna, el análisis de los factores extrabiológicos que afectan la biodiversidad, es una tarea no solo necesaria sino urgente. Sólo habría que señalar que México ocupa el tercer sitio en las tasas actuales de deforestación en el mundo, con una superficie anual estimada de 800000 has deforestadas (Massera, et al., 1992). Aquí de nuevo es necesario generar modelos de carácter multi-causal de los fenómenos destructivos (por ejemplo, la deforestación, véase Rotmans y Swart, 1991; Toledo, 1992).
Las dos fuentes principales de información para la construcción de estos modelos son: 1. Los acervos de información estadística o censal, que, a su vez, son el resultado de encuestas levantadas en el terreno, y que proporcionan información acerca de fenómenos demográficos, productivos, culturales, de tenencia de la tierra, de uso de energía, etcétera. 2. Las imágenes obtenidas de sistemas de percepción remota (satélites, fotografías aéreas, videos), y que ofrecen datos, a diferentes escalas espaciales, sobre los recursos naturales (vegetación, suelos, geología, topografía, hidrología, etcétera). El modo más adecuado y eficaz, para el manejo integrado de estos datos son los llamados Sistemas de Información Geográfica (SIG). Un SIG, “…es un conjunto de programas y equipos de computación que permite el acopio, manipulación y transformación de datos espaciales (mapas, imágenes de satélite) y no espaciales (atributos) provenientes de varias fuentes…”. El objetivo de un SIG es el de contribuir con información para la toma de decisiones y puede considerarse como una fuente para la conformación de sistemas expertos” (Bocco, et al., 1991:81). Estos sistemas, sin embargo, deben ser reconceptualizados desde una perspectiva ecológica, de tal suerte que sea posible la geo-referenciación de los factores socioeconómicos, en términos de los hábitats, que son los “escenarios teatrales” de la biodiversidad. Como es de esperarse, existe toda una gama de diagnósticos que pueden realizarse a través de estos sistemas. Habría que distinguir por lo menos dos tipos de diagnósticos: a. Los de carácter sincrónico, es decir, que ofrecen análisis para un tiempo determinado (por ejemplo, el presente); b. Los que analizan a través del tiempo y que permiten descubrir procesos diacrónicos o históricos (por ejemplo, las tendencias en la deforestación de una cierta área), mediante el empleo de imágenes y datos por ejemplo, censales) de diferentes épocas. En otra perspectiva también pueden plantearse modelos de carácter predictivo que buscarían realizar exámenes del futuro.
Agradecimientos
En la elaboración de este ensayo recibí oportunos apoyos de Ana Batis y Alejandro Torres en la búsqueda de información. Agradezco también a Carlos Vázquez-Yanes, John Fa, Claudio Delgadillo, Jorge Soberón, Victoria Sosa, Jorge González, Jorge Llorente, César Carrillo, Betty J. Meggers, Francisco Díaz-Pineda y Ma. de Jesús Ordóñez, por haberme facilitado materiales inéditos o de difícil acceso. La elaboración de varios cuadros y figuras fue posible gracias a la asistencia técnica de Leni Cortés.
|
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Referencias Bibliográficas
Ayala, R., T. R. Griswold y S. H. Bullock, 1993, “The native bees of Mexico”, en: Ramamoorthy, T. P., et al., 1993.
Bocco, G., J. L. Palacio y C. Valenzuela, 1991, Integración de la percepción remota y los sistemas de información geográfica, Ciencia y Desarrollo, 97:79-88. Brown, J. H, 1988, “Species Diversity”, en: Myers, A. A., P. S. Giller (eds.), Analytical Biogeography, Chapman and Hall Eds. :57-90. Cabrera, L., y J. L. Villaseñor, 1987, “Revisión bibliográfica sobre el conocimiento de la familia Compositae en México”, Biótica 12:131-141. Campbell, D. G., 1988, “The importance of floristic inventory in the tropics”, en: D. G. Campbell & H. D. Hammond (eds.), Floristic Inventory of Tropical Countries, New York Bot. Garden. Coates-Estrada, R., y A. Estrada, 1985, Lista de las Aves de la Estación de Biología de Los Tuxtlas, Instituto de Biología. UNAM. Delgadillo, C., 1987, “Moss distribution and the phytogeographical significance of the Neovolcanic Belt of Mexico”, Jour. Biogeography, 14: 69-78. Delgadillo, C., 1990, “Advances in Mexican Bryology”, Tropical Bryology, 2:49-52. Delgadillo, C., 1993, “Diversity in the Mexican Bryoflora”, en: Ramamoorthy, T. P., et al., (eds.): 365-377. Delgadillo, J. R., 1987, “Aspectos florísticos y ecológicos de las Sierras de Juárez y San Pedro Mártir, Baja California”, en: Resúmenes X Congreso Mexicano de Botánica, Soc. Bot. de México, 452. Di Castri, F. y T., Younes, (eds.), 1990, “Ecosystem Function of Biological Diversity”, Biology International, Special Issue, No 22. Erwin, T. L., 1991, “How many species are there?” Revisited, Conservation Biology, 5:330-333. Escalante, P. P, A. G. Navarro, A. T., y Peterson, 1993, “A Geographic, Ecological, and Historical Analysis of Land Bird Diversity in Mexico”, en Ramamoorthy, T. P., et al. (eds.) :281-307. Espinoza-Pérez, H. et al., 1993, “Notes on Mexican Ichthyofauna”, en: Ramamoorthy, T. P., et al. (eds.) :229-251. Fa, E. J., 1989, “Conservation-motivated analysis of Mammalian Biogeography in the Trans-Mexican Neovolcanic Belt”, National Geographic Reseach, 5:296-316. Fa, J., L. M. Morales, 1993, “Patterns of Mammalian Diversity in Mexico”, en: Ramamoorthy, T. P., et al., (eds.) :319-361. Field, C. B. and C. Vázquez-Yanes, 1993, “Species of the genus Piper provide a model to study how plants can grow in different kinds of rainforest habitats”, Interciencia (en prensa). Flores, O. y P. Gerez, 1989, Conservación en México: síntesis sobre vertebrados terrestres, vegetación y uso del suelo, INIREB/Conservation International. Gadow, H., 1907-1909, “Altitude and distribution of plants in Southern México”, Jour. Linn. Soc. Bot., 38: 429-440. García, E., 1989, Diversidad climático vegetal en México, trabajo presentado en el Simposio sobre Diversidad Biológica de México, Oaxtepec, Morelos, 18 pp. Gaston, K. J., 1991, “The magnitude of global insect species richness”, Conservation Biology, 5:283-296. Gentry, A. H., 1982, “Patterns of Neotropical plant species diversity”, Evolutionary Biology, 15: 1-84. Gentry, A. H., 1992, “Tropical forest biodiversity: distribution patterns and their conservation significance”, OIKOS, 63: 19-28. Halffter, G., 1976, “La entomofauna americana: ideas acerca de su origen y distribución”, Folia Entom. Mex., 6: 1-108. Kuhn, T., 1962, The Structure of Scientific Revolutions, Univ. Press Chicago. Léveque, F., M. Glachant, 1992, “Diversité Génétique. La gestion mondiale des ressources vivantes”, La Recherche, 23:116-123. Llorente-Bousquets J. y A. L. Martínez, “Conservation-Oriented Analysis of Mexican Butterflies: Papilionidae (Lepidoptera, Papilionoidea)”, en: Ramamoorthy, T. P., et al. (eds.), :147-177. Lot, A. y A. Novelo, P. Ramírez-García, 1993, “Diversity of Mexican Aquatic Vascular Plant Flora”, en: Ramamoorthy, T. P., et al., (eds.) :577- 591. Lovejoy, T. E. et al., 1986, “Edge and other effects of isolation of Amazon forest fragments”, en: Soulé, M. E. (ed.), Conservation Biology: the science of scarcity and diversity, Sinauer Assoc. Inc.: 257- 285. Mac Arthur, R. H., 1965, “Patterns in species diversity”, Biol. Rev., 40:510-533. Mac Arthur, R. H., 1972, Geographical Ecology, Harper & Row Pub., 269 pp. Magurran, A. E., 1988, Ecological diversity and its Measurement, Croom Helm, London. Margalef, R., 1957, La teoría de la información en ecología, Mem. Real Acad. Ciencias Barcelona, 32:373-449. Margalef, R., 1968, Perspectives in Ecological Theory, Univ. Of Chicago Press. Margalef, R., 1991, “Reflexiones sobre la diversidad y significado de su expresión cuantitativa”, en: Pineda, F. D., et al,. (eds.) Diversidad Biológica/Biological Diversity, Fundación Areces/WWF/SCOPE, Madrid: 105-112. Masera, O., M. J. Ordoñez & R. Dirzo, 1992, “Emisiones de carbono a partir de la deforestación en México”, Ciencia, 43: 151-153. May, R. M., 1992, “How many species inhabit the Earth?” Scientific American, October: 18-24. Maza, J. de la y R. G. de la Maza, 1985, “La fauna de mariposas de Boca del Chajul, Chiapas, México, (Rhopalocera)”. Parte 1. Rev. de la Soc. Mex. de Lepidopterología, 9:23-44. Mittermneir, R., 1990, “Wealth of plants and animals unites ‘megadiversity’ countries”, Tropicus, 4:4-5. McNeely, J. A., et al., 1990, Conserving the World’s Biological Diversity, IUCN, Gland Switzerland. Morales, J. C. & Llorente, J., 1986, “Estado actual del conocimiento de los siphonaptera de México”, An. Inst. Biol. Univ. Nal. Autón. Méx., 56:497-554. Pérez-Gil, R. et al., 1984, Ecosistemas de México, Banco BCH, México. Pielou, E. C., 1975, Ecological Diversity, Willey, NY. Pielou, E. C., 1991, “The many meanings of diversity”, en: Pineda, F. D., et al., (eds.) Diversidad Biológica/Biological Diversity, Fundación Areces/WWF/SCOPE, Madrid: 113-116. Pimentel, D., et al., 1992, “Conserving biological diversity in agricultural/forestry systems”, Bio-Science, 42:354-362. Ramamoorthy, T. P., R. Bye, A. Lot, J. Fa, (eds.) 1993, Biological Diversity of Mexico: Origins and Distribution, Oxford Univ. Press, New York, 812 pp. Ramírez-Pulido, J., C. Müdespacher, 1987, “Estado actual y perspectivas del conocimiento de los mamíferos de México”, Ciencias, 38:49-67. Rapoport, E., 1975, Aerografía: estrategias geográficas de las especies, FCE, México. Redford, et al., 1990, “There is more to biodiversity than the tropical rain forests”, Conservation Biology, 4:328-330. Rotmans, J. y R. J. Swart, 1991, “Modeling tropical deforestation and its consequences for global climate”, Ecological Modeling, 58:217-247. Rzedowski, J., 1978, Vegetación de México, Limusa, México. Rzedowski, J., 1993, “Diversity and Origins of the Phanerogamic Flora of Mexico”, en: Ramamoorthy, T. P., et al. (eds.) :129-144. Scott, J. M., 1987, et al., 1987, “Species richness: a geographic approach to protecting future biological diversity”, Bio-Science, 37:782-788. Shmida, A. y M. V. Wilson, 1985, “Biological determinants of species diversity”, Journal of Biogeography, 12:1-20. Soberón, J., y J. Llorente, 1993, “The use of species accumulation functions for the prediction of species richness”, Conservation Biology, (en prensa). Sosa, V. y P. Dávila, (en prensa), “Una evaluación del conocimiento florístico de México”, Ann. Missouri Bot. Garden. Soulé, M. E., 1985, “What is Conservation Biology”, Bio-Science, 35:727-734. Storck, N. y K. Gaston, 1990, “Counting species one by one”, New Scientist, August: 43-47. Toledo, V. M., 1982, “Pleistocene changes of vegetation in tropical Mexico”, en: G. Th. Prance (ed.), The Biological Diversification in the Tropics, Columbia Univ. Press: 93-111. Toledo, V. M., et al., 1985, Ecología y Autosuficiencia Alimentaria, Siglo XXI Eds., México. Toledo, V. M., 1988, La diversidad biológica de México, Ciencia y Desarrollo, 81: 17-30. Toledo, V. M., et al., 1989, La Producción Rural en México: alternativas ecológicas, Fundación Universo Veintiuno, México. Toledo, V. M., 1992, “Cambio climático y deforestación tropical: criterios para el análisis de un proceso complejo”, Ciencia, 43:129-134. Toledo, V. M., M. J. Ordóñez, 1993, “The Biodiversity Scenario of Mexico: A Review of Terrestrial Habitats”, en: Ramamoorthy, T. P., et al., (eds.), :757-777. Toledo, V. M., 1993, “La riqueza florística de México: un análisis para conservacionistas”, en: Guevara, S., P. Moreno (eds.), La Botánica Mexicana hacia el Fin del Milenio, Inst. de Ecología, (en prensa). Toledo, V. M., V. Sosa, 1993, “Floristics in Latin America and the Caribbean: an evaluation of the numbers of plant collections and botanists”, Taxon, 42:355-364. Toledo, V. M., & J. Rzedowski, 1993, “Floristics, vegetation, ethnofloristics and plant conservation in Mexico”, en: V. Heywood & O. Herrera MacBryde, (eds.) Centers of Plant Diversity: a guide and strategy for their conservation, Areces/WWF/SCOPE, (en prensa). Usher, M. B., 1991, “Biodiversity: a scientific challenge for resource managers in the 1990s”, en: F. D. Pineda, et al., (eds.), Diversidad Biológica/Biological Diversity, Fundación Areces/WWF/SCOPE, Madrid:33-40. West, R. C., 1964, The natural regions of Middle America, en: R. C. West (ed.) Handbook of Middle American Indians Vo1. 1, Univ. Of Texas Press: 363-383. Whittaker, R. H., 1972, “Evolution and measurement of species diversity”, Taxon 21:213-251. Wilson, E. O. (ed.), 1988, Biodiversity National Academic Press, Washington.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nota
Este texto es parte del libro: La diversidad biológica de México en los noventas, que será publicado próximamente por la Editorial Quinto Sol.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
____________________________________________________________
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Víctor Toledo
Centro de Ecología,
Universidad Nacional Autónoma de México.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ____________________________________________________________ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
cómo citar este artículo →
Toledo, Víctor Manuel. 1994. La diversidad biológica de México. Nuevos retos para la investigación en los noventas. Ciencias, núm. 34, abril-junio, pp. 42-59. [En línea].
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||





 |
|
||||||||||
| Jean Gayon | |||||||||||
|
A pesar de las numerosas críticas que se han expresado
a lo largo de los últimos dos decenios, el núcleo central de la teoría causal dominante de la evolución, apenas ha cambiado desde 1930. Esta teoría se basa fundamentalmente en dos proposiciones: la primera, afirma que la evolución es “un cambio en la composición genética de las poblaciones”,1 y la segunda, apoya la idea de que la selección natural es la fuerza mayor que orienta tal cambio.
Estas dos afirmaciones son uno de los retos mayores a los que se enfrenta la ontología espontánea de los biólogos, ontología que consiste, tradicionalmente, en considerar que los organismos individuales, —con su configuración espacial y su desarrollo— como verdaderas cosas sobre las cuales trata su ciencia. Ahora bien, si la biología de la evolución se interesa prioritariamente por entidades tales como los genes, las poblaciones y las especies, uno se asombra de que haya dicho tan poco en relación con la forma. En realidad, tales entidades figuran de manera paupérrima en el lenguaje de la forma, sea cual fuere el sentido que se le dé a este término.
El objeto de este texto es demostrar cómo la visión moderna de la especie biológica y de su evolución se ha formado pagando el precio de una marginalización cada vez más profunda del concepto de forma; el evolucionismo darwiniano y mendeliano fue, de hecho, el que logró una antigua empresa que era la de hacer del concepto de especie, un concepto despojado de toda alusión a la forma y, en consecuencia, al significado original, propiamente dicho, de la palabra especie.
El concepto de especie
Si queremos destacar las dificultades producidas por el concepto moderno de especie, nos será de utilidad recordar la herencia semántica de esta palabra. Especie (species) es la traducción latina del griego eïdos. Sin embargo, existe otra traducción filosófica convencional de eïdos: es la de “forma”. En la lengua griega, la formación de la constelación semántica del término eïdos puede resumirse en las etapas siguientes.2 Todo sucede en realidad en la lengua preplatónica. La significación conocida más antigua es la que se refiere a una abstracción en relación con la faz humana, que en seguida se extendió a la designación del aspecto visible del cuerpo, uso que los propios médicos griegos radicalizaron cuando se sirvieron del eïdos para referirse a la constitución interna del cuerpo. En una generalización posterior, eïdos designó la forma espacial en general, en razón de la proximidad etimológica con idea, que designaba la forma geométrica abstracta de cierta cosa (por ejemplo, la forma de los átomos en Demócrito). Simultáneamente, Hipócrates impuso el uso de eïdos como tipo constitucional, uso que se generalizaría una última vez entre los primeros filósofos y que desembocaría en la noción categórica de eïdos como clase lógica. Vemos entonces bastante bien la compleja constelación semántica de eïdos y lo cercana que se encontró de un concepto de representación del cuerpo vivo.
El corpus aristotélico muestra, de hecho, cómo el discurso de la vida se volvió, desde temprano, una teoría en favor de una tensión entre numerosos conceptos de eïdos. En Aristóteles, compiten fundamentalmente dos conceptos de eïdos en la representación de lo vivo. Uno de ellos es el de clase lógica: utilizado en unión con el de género (genos), se aplica en derecho a todos los dominios de la realidad y constituye un instrumento de jerarquización de los universales. Este concepto de eïdos como clase lógica, se aplica sin restricciones a los seres vivos, sobre todo cuando se trata precisamente de clases. Sin embargo, los tratados biológicos muestran otro concepto, el de eïdos como alma del organismo individual, aquel por medio del cual Aristóteles designa un principio organizador transmisible por medio de la generación.
A diferencia del primer sentido de eïdos, el eïdos como universal, esta segunda acepción envuelve connotaciones espaciales y temporales (estructura [schèma], forma en desarrollo [ruthmos] y forma generativa [morphè]). Finalmente, en ciertos textos raros y extraños (Generación de los animales, 731a 31-732b 1), Aristóteles utiliza eïdos en un sentido que subordina el eïdos lógico al eïdos generativo. Nos dice, en efecto, que los seres vivos, si no son individualmente eternos, lo son sin embargo “por el eïdos” transmitido en la generación, y que es en virtud de este proceso que existen “géneros” entre los hombres, los animales y las plantas. La subordinación ordinaria del género y de la especie se encuentra así invertida; Aristóteles parece admitir, al menos en un texto, que la coherencia propia de las entidades taxonómicas estaría fundada en la naturaleza de la continuidad genealógica.
Vemos entonces cómo el estatuto ontológico de la especie constituyó desde temprana época una pregunta comprometedora para los naturalistas. Tal ha sido el primigenio momento de la historia del concepto biológico de especie, historia que ha consistido en gran parte en marginar cada vez más las connotaciones de la clase lógica y las de forma; en consecuencia, de las dos significaciones principales y concurrentes del eïdos en la lengua culta de los griegos. Enseguida, resumiré tres etapas cruciales de esta historia.
El primer momento, conceptualmente decisivo, debe ser buscado en la célebre controversia preevolucionista de Buffon con Linneo. Contra Linneo, que consideraba que cada especie era identificable a través de una marca morfológica unívoca, Buffon puso de relieve un nuevo concepto de especie, en ruptura con su representación escolástica tradicional como un “universal” o “clases” bien definido de individuo. Para Buffon, la especie no tiene el estatuto absoluto de una clase intemporal sino el de una cosa física cuya coherencia está definida por la sucesión genealógica.
Existe un bello texto que expresa claramente el paso del primer al segundo concepto. Se trata de un pasaje del artículo de la Histoire naturelle dedicado, en 1753, al “Asno”:
“un individuo es un ser aparte, aislado, separado, y que no tiene nada en común con todos los otros seres, si no fuera por el hecho de que se les parece o bien que de ellos difiere: todos los individuos semejantes que existen sobre la superficie de la Tierra son vistos como componentes de la especie de individuos” (subrayado por mí).
En esta primera fase, “especie” es tomada en el sentido filosófico tradicional de clase lógica de objetos (“una especie de”, “una clase de”). En seguida, Buffon escribe:
“sin embargo, no es ni el número ni la colección de los individuos semejantes lo que hace a la especie, es la sucesión constante, así como la renovación de estos individuos que la constituyen (…) la comparación del número y de la semejanza de los individuos no es más que una idea accesoria y a menudo independiente de la primera”.3
Buffon postula entonces que, entre los seres vivos, la relación de semejanza debe estar subordinada a aquella de sucesión en la generación. En efecto, señala con insistencia, que de todos los criterios utilizados intencionalmente por los taxonomistas para definir la categoría de especie, ninguno es verdaderamente universal: ni la morfología, ni las condiciones de vida, ni el comportamiento, ni siquiera el aislamiento reproductivo, permiten definir de manera unívoca a la categoría de especie. Así, el naturalista debe reconocer la absoluta prioridad de la continuidad de la descendencia en la definición de la categoría de especie viva.
Con Buffon, nos encontramos de hecho ante un giro mayor de la historia del concepto de especie, puesto que con él se trata de reservar el concepto de especie a este grupo de seres naturales que exigen que uno subordine la clasificación a la genealogía. Buffon, de manera muy coherente, invita a los científicos a no utilizar el término de especie para los minerales; por ejemplo, esta reflexión de Buffon sobre las especies como entidades genealógicas es aún más notable cuanto que se produjo en un contexto preevolucionista.
La segunda etapa, consiste en la formación del concepto moderno de especie biológica, tal como se ligaría precisamente con el transformismo del siglo XIX. Mientras que Buffon había emancipado a las especies biológicas del estatuto de universales, el pensamiento naturalista del siglo XIX marginó al otro aspecto del eïdos, el de forma. No quiero decir aquí que la morfología haya sido despreciable en la edificación de la visión evolucionista de la historia de la vida. Con toda seguridad, fueron numerosos los morfólogos del siglo XIX que pensaron poder dar cuenta de la sucesión geológica de las forma vivas por medio de una lógica de las formas. Los naturalistas alemanes de la escuela de Göttingen (Blumenbach, Treviranus), los trascendentalistas alemanes y después franceses (Goethe, Geoffroy St.-Hilaire, etcétera), Owen en Inglaterra, Agassiz en Suiza y después en los Estados Unidos, se adhirieron a esta idea, fundada sobre un método de ordenación de las formas por medio de la transformación topológica de un plan de organización fundamental.
Sin embargo, este método entró en conflicto con otra imagen del orden natural, desarrollada al mismo tiempo por aquellos que estudiaban las formas vivas en su contexto geográfico. Los fundadores de la fitogeografía (A. von Humboldt, A. P. de Candolle) atrajeron la atención de los naturalistas sobre las similitudes entre los seres vivos en razón de su hábitat. De ahí proviene la idea de una tipología funcional de las formas, muy diferente de la tipología de los morfologistas trascendentalistas, más interesados por los problemas de las servidumbres topológicas. Los biogeógrafos insistieron, entre otras cosas, sobre el hecho de que las estructuras anatómicas recientes semejantes, podían ser asociadas con especializaciones adaptativas extremadamente diferentes (por ejemplo, el miembro tetrápodo: a veces una pata, a veces una ala, a veces una aleta). Pero, al mismo tiempo, los primeros biogeógrafos se encontraban, con razón, con el hecho enigmático de que tantas especies, con sus especializaciones adaptativas propias, estuvieran confinadas en áreas geográficas estrechas, a pesar de la existencia de hábitats semejantes en otras partes del mundo: una tipología puramente funcional ya no era entonces suficiente.
Uno de los méritos mayores de Darwin consistió en haber propuesto una interpretación teórica coherente de estas paradojas.4 Éstas se pueden reducir en dos proposiciones sencillas; la primera es una respuesta directa al problema del desacuerdo entre estructuras profundas y especializaciones adaptativas: las estructuras profundas son las trazas de un pasado lejano de la especie; las especializaciones adaptativas deben comprenderse a partir de las condiciones de vida actuales. Dos inteligibilidades se sobreponen entonces a dos escalas temporales diferentes: aquella de la descendencia (o “herencia”) y aquella de las adaptaciones, siempre relativas.
El segundo pilar de la visión darwiniana de las especies, se encuentra en el postulado de la universalidad de la variación: esta es la norma para todos los caracteres, en todas las especies y en todo momento. El postulado es universal para garantizar la eficacia del proceso de selección natural, base fundamental de la teoría de la descendencia con modificación. En efecto, si todas las especies presentan continuamente una variación heredable por un gran número de caracteres, siempre hay lugar para una transformación oportunista de la especie.
En consecuencia, con Darwin, la meta ontológica de la explicación naturalista se desplaza de la forma hacia la especie. La especie darwiniana no es una forma: es un grupo de individuos variables cuya coherencia propia es principalmente reproductiva. Este grupo reproductivo es, propiamente dicho, la cosa física que se modifica: esta cosa no tiene el estatuto de una clase lógica; solo una cosa, no un abstracto, puede cambiar.
En cuanto a la forma en el sentido de configuración, no es un atributo de la especie propiamente dicha, sino de los organismos individuales que componen a la especie. Es de hecho extremadamente difícil concebir a una especie biológica darwiniana en términos de organización espacial. De todas las entidades biológicas reales, las especies darwinianas son probablemente aquellas que tienen el más bajo grado de organización; no por ello dejan, sin embargo, de ser el campo fenoménico al interior del cual se encuentra controlado el cambio evolutivo. Tenemos ahí una situación epistemológica en verdad bastante extraña. Muchos biólogos y filósofos se han equivocado en relación con la implicación filosófica de la visión darwiniana del origen de las especies para la comprensión de la historia de la vida. Darwin no ha construido una teoría de la génesis de las formas, aunque a veces haya sugerido una interpretación de su obra en semejantes términos. El darwinismo no es una metodología capaz de decirnos cuáles formas son posibles y cuáles son las leyes susceptibles de dar cuenta de la génesis de tal o tal forma; es una hipótesis sobre las condiciones más generales bajo las cuales las formas vivas van a ser localmente modificadas. De ahí el estatus muy particular del principio de selección natural, que nunca es representable más que en referencia a una acumulación de contingencias locales (a diferencia de los grandes principios físicos, la selección natural no es una “ley-origen”, sino una “ley-consecuencia”).
Nos queda un último paso a dar para concebir plenamente la marginación de la forma en la biología evolutiva moderna. En la primera mitad del siglo XX, la teoría darwiniana se encontró reformulada en el vocabulario de la genética y de la biología de las poblaciones. En este nuevo contexto, aun más que en Darwin, el proceso evolutivo no afecta a los organismos individuales; afecta a las poblaciones mendelianas en el seno de las cuales los genes se encuentran muestreados, combinados, mutados, seleccionados. (El paradigma de Dobzhansky de 1937: evolution is a change in the genetic composition of population).
No quiero aquí más que señalar las implicaciones de semejante paradigma de investigación en la perspectiva de una reflexión sobre la forma biológica. Al hacer de la genética de las poblaciones el núcleo de la teoría causal de la evolución, la biología moderna ha escogido privilegiar aquellos sujetos cuya significación morfológica es la más tenue. La cosa es bastante evidente en relación con las poblaciones; lo es todavía más para las especies, que no son más que reservas de poblaciones virtualmente interfecundas. Pero esto es verdadero también para los genes. Por supuesto, sabemos desde hace 40 años que los genes tienen una composición y una conformación molecular definidas rígidamente. Sin embargo, en la cadena causal que conduce a la forma observable de un individuo, los genes ocupan una posición muy alejada. Por eso, la genética de las poblaciones aparece como una ciencia que no tiene nada que decir sobre la forma de los individuos, aunque sea la teoría central de la biología causal de la evolución. Para utilizar una expresión de Lotka de los años 1920, la genética de las poblaciones es una pura cinemática formal, que hace abstracción de las cualidades de los objetos que evolucionan, y que se contenta con describir su ritmo evolutivo, bajo ciertas condiciones específicas.5
Llegamos así al término de una empresa que ha pretendido dejar de lado a la forma en las teorías biológicas, aquella de la teoría causal de la evolución que pretende aun más que todas las demás, dar cuenta de la unidad profunda de las ciencias de la vida. Nacida hace cerca de dos siglos, la ciencia de la modificación de las especies se ha liberado poco a poco de la inteligibilidad geométrica y se ha construido alrededor de entidades —los genes, la población, la especie— que eliminan toda referencia (o casi) a la forma aparente de los organismos. Al mismo tiempo, la teoría de la descendencia ha terminado por especificar al concepto de especie como tipo, o clase homogénea de individuos. Me parece entonces bastante claro que la biología de la evolución se ha orientado cada vez más, hasta una época reciente, hacia un divorcio entre las significaciones diversas de eso que los griegos llamaban eïdos: ni tipo, ni forma, la especie darwiniana no se concibe más que como una singularidad histórica, dotada sin duda de una fabulosa memoria, pero una memoria que no podemos comprender más que de manera local.
Esto dibuja sensu contrario el perfil de lo que podría convertirse en la biología futura de la evolución; hay que esperar que ella se desplace, en relación con su futuro, hacia un nivel de comprensión más general y que se interese menos en las condiciones de posibilidad del cambio local. Por el contrario, se interesará más en la descripción de lo posible: es decir, en saber cuáles son los límites de lo posible en materia de organización del genoma, en materia de programación de desarrollo o en materia de estrategia ecológica a la escala de la biósfera y de los tiempos geológicos. El filósofo no puede aquí más que registrar las tendencias manifiestas que observa en el campo de la embriología, de la ecología y de la dinámica de los sistemas complejos. Si semejantes tendencias se confirmaran, podríamos apostar que la futura teoría de la evolución sería tanto más una teoría sobre el origen de la vida en grande (macrovida) que una teoría sobre el origen de las especies. Pero es muy temprano para que el búho de Minerva emprenda su vuelo sobre este paisaje.6
Agradecimientos
El autor agradece a Jorge Martínez Contreras, profesor de la Universidad Autónoma Metropolitana Unidad Iztapalapa, el generoso apoyo en la organización de la mesa redonda “El darwinismo hoy”, realizada en la Universidad Autónoma Metropolitana Unidad Xochimilco, el 28 de octubre de 1993.
|
|
|
|
||||||||
|
Nota
Traducción de Jorge Martínez Contreras
|
|||||||||||
|
Referencias Bibliográficas
1. Dobzhansky, Th., Genetics and the Origin of Species, New York, Columbia University Press, p. 11.
2. Sandoz, Charles, 1971, Les noms grecs de la forme, tesis, Neuchatel. 3. Buffon, G. Leclerd de, Histoire naturelle, t. III [1753], L’asne. 4. Canguilhem, G., et al., Cf., Du développement à l’évolution, Paris, Presses Universitaires de France. 5. Gayon, J., Cf., 1992, Darwin et l’après-Darwin: une histoire de l’hypothèse de sélection naturelle, Paris, Editions Kimé, Chap. IX. 6. Para una versión desarrollada de este texto, ver: “L’espèce sans la forme”, en J. Gayon & J. J. Wunenburger, eds. 1992, Les figures de la forme, Paris, L’Harmattan, pp. 49-61. También: “L’individualité de l’espèce: une thèse transformiste?”, en J. Gayon, ed., Buffon 88, Paris, Vrin, 1992, pp. 475-489. |
|||||||||||
| __________________________________ | |||||||||||
|
Jean Gayon
Universidad de Borgoña, Francia
|
|||||||||||
| ____________________________________________________ | |||||||||||
|
cómo citar este artículo →
Gayon, Jean. 1994. La marginación de la forma en la biología darwiniana. Ciencias, núm. 34, abril-junio, pp. 30-34. [En línea].
|
|||||||||||
 |
|
||||||||||
| Enrique Forero | |||||||||||
|
Sin lugar a dudas la flora tropical es la más rica del mundo
en diversidad vegetal pero también es la menos conocida. Del total de plantas vasculares (se calcula que en el mundo hay cerca de 250000 especies), se cree que cerca de 150000 crecen en las regiones tropicales. Otros grupos importantes incluyen unos 11000 helechos y grupos afines y 16000 briofitos. África tropical tiene el menor número de especies de angiospermas, con cerca de 30000. En Asia tropical, Australia y la región del Pacífico hay por lo menos 35000 especies. La diversidad biológica de América tropical, por otra parte, no tiene igual. La flora de América Latina es la más rica del mundo en número de especies, pues alcanza probablemente las 90000 (o sea cerca del 38%).
Además de su considerable diversidad de especies, la región neotropical es también muy rica en diversidad de hábitats. Las selvas más conocidas son las de las tierras bajas de la Amazonia o de la costa del Pacífico, en la región biogeográfica del Chocó. Existen, sin embargo, franjas altitudinales de vegetación con gran cantidad de elementos f1orísticos provenientes de las zonas templadas del norte y del sur, y una excepcional flora de altas montañas o “alpina”, como la de los páramos de América del Sur; asimismo, se encuentran zonas áridas con desiertos o matorrales, sabanas y cerrados, y muchos otros tipos diferentes de bosques. Esta diversidad de hábitats tiene gran importancia para la riqueza y diversidad de la flora de la región.
En un artículo publicado en 1977, haciendo referencia a la riqueza florística de América Latina, Prance escribió lo siguiente: “La tragedia del inventario biológico en los trópicos es que la destrucción de la vegetación avanza más rápido que el inventario”. La deforestación, como todos sabemos, depende de varios factores asociados con los problemas económicos y sociales que existen en los países en desarrollo. El crecimiento de la población, la presión sobre los recursos naturales disponibles, la ganadería, el desarrollo industrial, la construcción de carreteras y caminos, las concesiones madereras, la mala administración y la deficiente planificación, y la falta de estudios de impacto ambiental han contribuido en forma considerable a este proceso de destrucción.
De acuerdo con Henderson, Churchill y Luteyn, en la región Amazónica con un área de 7500000 km2 y una riqueza florística calculada en 30000 especies, se ha perdido entre el 8 y el 11% de la superficie forestal total, mientras que en el extremo norte de los Andes —un área de 383000 km2 y una flora calculada en cerca de 40000 especies— la superficie deforestada alcanza al 90 o 95%. La situación en los bosques de la costa Atlántica del Brasil es similar: con un área de 1000000 de km2 y una flora calculada en unas 10000 especies, el área deforestada alcanza un 95%.
Los acuerdos de Río
En junio de 1992, 178 países se dieron cita en Río de Janeiro en la reunión cumbre de mayor envergadura realizada en el mundo hasta ese momento, la Conferencia de las Naciones Unidas sobre el Medio Ambiente y el Desarrollo, también conocida como “Cumbre de la Tierra”.
Su tarea era, como lo señala Robinson, la de guiar a la humanidad hacia el llamado “desarrollo sostenido”. El concepto general era el de integrar valores ambientales y conceptos económicos, de manera que la humanidad pueda responder a sus necesidades actuales (equidad intrageneracional) sin poner en peligro la posibilidad de las futuras generaciones de responder a sus propias necesidades (equidad intergeneracional).
Motivada por el interés en buscar una solución a esos problemas, la Asamblea General de las Naciones Unidas estableció una Comisión Preparatoria para la Conferencia de Río-92 y, al mismo tiempo, comenzó las negociaciones internacionales para preparar las convenciones sobre cambios climáticos y sobre biodiversidad.
Durante la Conferencia en Río de Janeiro, la Convención Marco sobre cambios climáticos, fue firmada por 154 países y la Convención sobre Diversidad Biológica fue firmada por 156.
Asimismo, se adoptaron la Declaración de Río sobre el medio ambiente y el desarrollo y la Agenda 21 (ver recuadros).
Las realidades del mundo en desarrollo El logro de muchos de los objetivos de las convenciones o de Agenda 21 depende de que se realicen profundas reformas en muchos aspectos de la situación mundial actual.
Uno de los problemas fundamentales que enfrentan los países en vías de desarrollo es la deuda externa. El pago de la deuda externa es equivalente a una transfusión de sangre de una persona enferma a una persona saludable. En 1972, los países en desarrollo debían aproximadamente 50000 millones de dólares a los países industrializados. En 1981 esa cantidad había crecido a 315000, Y para 1991 era de 445000 millones de dólares. La transferencia de capitales por año de los países del Tercer Mundo a los países industrializados es de 40000 millones de dólares, mientras el norte envía en asistencia entre 30000 y 40000 millones por año.
El Programa de las Naciones Unidas para el Desarrollo (PNUD), el Banco Mundial y los bancos de desarrollo regional, lo mismo que otras agencias, tienen que reorganizar sus prioridades para cumplir con los objetivos de Agenda 21 y de las convenciones. El GEF (Global Environmental Facility) ha sido creado bajo los auspicios del Banco Mundial como el vehículo para financiar las dos convenciones, la de biodiversidad y la de clima, lo mismo que otros proyectos de protección ambiental. Es necesario reemplazar los sistemas del pasado que produjeron un mundo lleno de deudas con un proceso más transparente que incluya a todos los interesados. El GEF, sin embargo, ha sido duramente criticado por diversos grupos que no lo ven como el mecanismo adecuado para financiar los proyectos en pequeña escala, necesarios para alcanzar la conservación de la biodiversidad y el uso sostenido de sus componentes en todo el mundo, y piensan que el trabajo del GEF debe abrirse a la participación de otras organizaciones (véanse por ejemplo, Caribbean Conservation News Vol. 5, No. 19, septiembre de 1992; “The Biodiversity Coalition”, Newsletter No. 5, 5 de junio de 1993; Biology International No. 27, julio de 1993).
Otro problema de grandes proporciones para el desarrollo científico de los países del tercer mundo es el acceso desigual a la información, al conocimiento y a las comunicaciones. En esos países la distribución desigual de la capacidad de aprender, saber y comunicarse es una de las principales fuentes de frustración de la comunidad científica.
El fortalecimiento de la comunidad científica, el asumir responsabilidades tanto por parte de la comunidad como de cada uno de sus componentes, y la producción y transferencia de información pertinente dentro y fuera de la comunidad y entre ésta y otras comunidades similares, son condiciones fundamentales para un proceso de desarrollo participativo y, eventualmente, para un desarrollo científico coherente a nivel mundial.
Los modelos de desarrollo científico estándar, importados de otras partes del mundo, son un obstáculo para la erradicación del subdesarrollo científico, porque están basados en criterios que no necesariamente son aplicables en todo el mundo. De acuerdo con esos modelos, el crecimiento en productividad científica depende de un proceso de producción altamente tecnificado, y el principal obstáculo para alcanzar el desarrollo continúa siendo la carencia de dinero. Queda claro que abandonar ciertas tecnologías nuevas como la biotecnología o la informática sería absurdo. Pero es importante combinarlas con otras tecnologías, ya sean tradicionales o de otros tipos. Existe un conocimiento técnico popular con el cual podría obtenerse un considerable beneficio social. Tenemos que aprender a experimentar, comparar, transferir y, de ser posible, generalizar experiencias exitosas. En cuanto a la carencia de dinero, todos sabemos que las transferencias de fondos mal concebidas, casi siempre generan efectos negativos; a menudo se usan para cosas diferentes de las que se pretendían originalmente, acentúan la desigualdad y la dependencia, penalizan la producción y dan lugar a prácticas corruptas.
No hay duda de que las comunidades científicas de los países en desarrollo requieren información y tecnología de frontera. Pero ese tipo de apoyo sólo funciona con la aceptación, paralelamente, del pluralismo, de una cierta medida de equidad social y de la responsabilidad que deben asumir las comunidades y cada uno de los individuos que las forman.
Si existen una organización colectiva y una transferencia de tecnología mejores, las cosas funcionarían también de mejor manera, combinando más correctamente las iniciativas individuales y colectivas, elevando la calidad de la producción científica, reduciendo costos, y manejando adecuadamente las condiciones de trabajo.
Finalmente, un análisis real de las condiciones en que se desenvuelve la comunidad científica en los países del tercer mundo, y un oído atento para escuchar y entender la forma en que ellos perciben su ambiente diario y las mejoras que podrían efectuarse rápidamente, constituyen, sin duda, fuentes importantes de progreso. Está demostrado que si se cuenta con medios adecuados de comunicación, información y organización propia, estas comunidades son capaces de encontrar las soluciones a sus dificultades.
La botánica en América Latina
¿Cómo se reflejan los asuntos tratados hasta ahora en la actividad botánica en América Latina?
Tasa de colecciones botánicas
Prance y Campbell han analizado los índices de colección de material de herbario en diversas regiones del mundo. Tomando como base el análisis de cada edición del Index Herbariorum de 1952 a 1981, y calculando cuántos especímenes por cada 100 km2 por año se habían agregado a los herbarios locales, estos autores encuentran que el inventario florístico en México y en la mayor parte de América Central ha aumentado desde 1974.
La situación en Belice, que aparecía como una excepción en ese análisis, ha mejorado en años recientes gracias a los esfuerzos de colección que han realizado el New York Botanical Garden y otras instituciones; mientras tanto El Salvador y Nicaragua han sufrido considerablemente debido a la difícil situación política que han vivido en años recientes. Los altos índices que registra Jamaica, deben ser analizados con cuidado pues, de acuerdo con Tom Zanoni, no se han realizado colecciones importantes en ese país desde 1980, y es posible que muchas de las colecciones que están depositadas en el herbario del Institute of Jamaica no sean plantas nativas de ese país. Zanoni también señala que la mayor parte del crecimiento en la tasa de colección en la República Dominicana durante los últimos trece años se debe al trabajo de los investigadores del Jardín Botánico de Santo Domingo.
Antes de 1981, los índices de colección también habían aumentado en América del Sur, excepto en las Guayanas, pero continuaban siendo muy bajos en comparación con México y con la mayor parte de América Central y el Caribe. Por ejemplo, el índice más alto era el de Colombia, con 1.8 ejemplares por 100 km2 por año, que es mucho menor a los de casi toda América Central (El Salvador aparecía con 24 ejemplares por 100 km2 por año). De acuerdo con estos autores, se requerirían cerca de 39 años para que en Colombia se acumularan 100 ejemplares por 100 km2 por año, lo cual se considera como un nivel mínimo para un inventario adecuado. Toledo y Sosa dieron información de índices de colección para América Latina y el Caribe, basándose en datos de 1990.
Herbarios, ejemplares y taxónomos
Los datos anteriores reflejan la actividad de los botánicos y de los herbarios locales para realizar el inventario de la flora de América Latina. Toledo y Sosa presentaron información sobre herbarios, especímenes y botánicos de la región. Compararon datos correspondientes a los años 1974, 1981 Y 1990. El número de herbarios en México aumentó de 18 a 71 desde 1974, pero en los demás países de América Central y del Caribe no ha habido cambios notables. En cuanto a América del Sur, sí se registró aumento en el número de herbarios en todos los países y, para 1990, ya había al menos tres herbarios activos en Bolivia.
Toledo y Sosa también calcularon que existen 2637921 ejemplares de herbario en México y América Central, 1008616 ejemplares en el Caribe, y 9710858 en América del Sur, lo que da un total de 13357395 en toda América Latina.
El número de botánicos que trabajan en América Latina y el Caribe ha aumentado en los últimos veinte años en casi todos los países, principalmente en Brasil, Perú, Costa Rica, Bolivia y Paraguay. Hoy alcanza un total de 1344 en toda la región. En Bolivia y Paraguay el inventario florístico se detuvo por muchos años. Afortunadamente, a partir de 1980 se crearon nuevas instituciones botánicas, y actualmente se cuenta con grupos de jóvenes botánicos muy capacitados, que están contribuyendo al estudio de la flora.
Un futuro incierto
¿Qué se puede hacer? ¿En dónde estamos y hacia dónde vamos? ¿Qué acciones concretas se pueden llevar a cabo para resolver los principales problemas que limitan el desarrollo de la ciencia botánica en México y en otros países en desarrollo, ante las nuevas condiciones mundiales?
Fortalecimiento de la comunidad científica
Es preocupante el hecho de que muchas de las instituciones que hasta ahora han sido las encargadas de estudiar la biodiversidad en gran parte de los países, no estén recibiendo el apoyo gubernamental e internacional que necesitan para continuar desarrollando esas funciones.
Es también preocupante que muchos jóvenes científicos latinoamericanos muy inteligentes y bien capacitados para hacer contribuciones importantes a la solución de los problemas ambientales que afectan a la región, estén subempleados o desempleados, y que muchos están abandonando sus países de origen o la profesión misma para dedicarse a otras actividades, en busca de mejores horizontes.
El papel de la comunidad científica
La comunidad académica tiene que tomar una nueva actitud y volverse más agresiva. El mundo moderno presenta una serie de retos que no pueden ser ignorados.
Es necesario aumentar la conciencia de que la misión de la ciencia y de la tecnología no es solo descubrir los hechos sino también la forma de aplicar esos nuevos conocimientos.
Es necesaria una mejor organización colectiva para poder enfrentar los retos del mundo moderno. Les compete a las sociedades botánicas de los distintos países, organizar grupos de trabajo que discutan muchos de los problemas que afectan a la comunidad en general, y que van desde el cómo actuar ante los gobiernos, las organizaciones no gubernamentales, las entidades financieras, etcétera, hasta el cómo participar en los procesos de decisión, tanto a nivel científico como de distribución de recursos, transferencia de tecnología, modernización, decisiones de la Conferencia de Río, establecimiento de prioridades en investigación, conservación, desarrollo sostenido, etcétera.
Como individuos, los botánicos deben preguntarse qué están haciendo en bien de la comunidad, no solo científica sino de la comunidad en general, de los ciudadanos. Cada uno debe asumir la responsabilidad que le corresponde.
México tiene la capacidad de asumir una posición de liderazgo en el concierto latinoamericano dada su experiencia, la importancia de sus instituciones científicas y, sobre todo, la alta calidad de su comunidad científica.
Producción y transferencia de información
Los científicos de los países de América Latina deben trabajar en colaboración con los científicos de otros países para agilizar el proceso de estudio de la biodiversidad y para producir la información que requieren tanto el público en general, como las entidades que toman decisiones.
Es necesario elaborar bases de datos a varios niveles, y establecer redes computarizadas de información.
También es indispensable compartir la información y el conocimiento con los colegas de otros países de la región. La Red Latinoamericana de Botánica es un buen ejemplo de colaboración y coordinación. En ella México ha jugado un papel muy importante.
No importa lo que se haya avanzado, todavía hay que buscar el camino para incrementar el intercambio de conocimientos y experiencias. Por eso la asistencia a reuniones científicas, dentro y fuera de la región requiere de mayor apoyo financiero. La participación de científicos de América Latina en reuniones, bien sea de carácter nacional, regional o mundial es un verdadero via crucis, lo que da como resultado atraso, falta de información y aislamiento.
La comunidad científica tiene que encontrar las formas más adecuadas y equitativas de colaboración con las organizaciones no gubernamentales. Cada una tiene algo que ofrecer a la otra, pero esta colaboración es aún difícil.
Asimismo, la comunidad científica tiene que esforzarse por hacer llegar al público una información útil, clara, y científicamente correcta.
Modelos importados de desarrollo científico
Los países de América Latina tienen que definir sus propios modelos de desarrollo científico. Como ocurre con muchos modelos ecológicos, las ideas venidas del norte, basadas en condiciones diferentes, no necesariamente son aplicables de manera directa a la realidad latinoamericana. Por eso es oportuno recapacitar cuidadosamente sobre la forma en que se aceptan los modelos que vienen de fuera. Esto incluye, pero no se restringe, a los métodos de evaluación del trabajo científico y, por consiguiente, a los métodos de financiación, a los modelos de estudio y conservación de la biodiversidad, a los modelos de coordinación interinstitucional, etcétera.
Transferencia de fondos
Las agencias financieras, tanto nacionales como internacionales, tienen que aceptar su responsabilidad en cuanto a apoyar el estudio, conservación y desarrollo sostenido de los recursos naturales. Esto incluye, como elementos muy importantes, el fortalecimiento de centros de investigación y el entrenamiento de nuevas generaciones de científicos.
La investigación taxonómica básica, los inventarios, las colecciones de germoplasma, las investigaciones etnobotánicas y, en general, las investigaciones de campo, deben recibir todo el apoyo posible, porque sólo así se podrá obtener la información necesaria para tomar decisiones inteligentes en relación con la protección, el manejo y el uso de los recursos vegetales de la región.
El Consejo Nacional de la Flora de México es la entidad apropiada para completar el inventario florístico del país. Pero, para ello, requiere de un apoyo financiero decidido.
Asimismo, los jardines botánicos y las instituciones regionales deben contar con un apoyo para realizar su importante contribución científica y educativa a diversos niveles.
Pluralismo y conocimiento local
Cualquier solución a los problemas ambientales tiene que ir acompañada de una participación activa de la comunidad que se verá directamente afectada por esas soluciones, por lo que éstas tienen que ser compatibles con las condiciones locales tanto de índole científica y técnica, como social y económica.
La combinación de tecnologías modernas como la biotecnología, con estudios sobre tecnologías tradicionales que se realizan a través de investigaciones etnobotánicas o de botánica económica, permitirá obtener soluciones locales enfocadas al mantenimiento de la biodiversidad.
Se habla mucho de desarrollo sostenido, pero éste no se alcanzará si no se tienen los conocimientos fundamentales de campo, de qué es lo que se quiere desarrollar en una forma sostenida.
Acuerdos, realidades y perspectivas
Todo lo anterior solo se logrará después de hacer un análisis cuidadoso de las condiciones reales en que se desenvuelven los países de la región: condiciones sociales, económicas, políticas y de ciencia y tecnología.
Las decisiones de la Conferencia de Río sólo podrán aplicarse en formas que sean convenientes y aceptables para los países en desarrollo, y en particular para los países tropicales, después de que éstos acepten la responsabilidad de decidir cuáles son sus propias prioridades y no necesariamente las prioridades que la comunidad internacional les presenta, y que con frecuencia les impone. La comunidad internacional debe estar dispuesta a aceptar esas prioridades y a proveer el apoyo tecnológico, científico y financiero necesario y requerido por los países.
La investigación científica tiene que convertirse en un pilar fundamental de la administración ambiental. Toda decisión que tenga que ver con biodiversidad en América Latina, debe tomar en cuenta lo que se está haciendo y también lo que no se está haciendo en la región. Todas esas decisiones tienen que tomarse contando con la activa participación de la comunidad científica local. Por esta razón, las organizaciones internacionales de toda índole, en todas sus deliberaciones, deben tomar la decisión real de involucrar, a representantes de los países poseedores de la mayor diversidad biológica del mundo.
En 1994, del 2 al 8 de octubre, se realizará en Mar del Plata, Argentina, el VI Congreso Latinoamericano de Botánica. Este evento continúa una tradición iniciada en México en 1972, y deberá constituirse en un momento histórico para la ciencia botánica en América Latina.
La comunidad botánica de la región tiene que prepararse para llegar al congreso a discutir en detalle los temas abordados en esta contribución. Hay tiempo suficiente para meditar sobre estos asuntos, sobre la participación de la comunidad científica en las decisiones de Río 92 y sobre el futuro de América Latina como poseedora de la mayor diversidad biológica del planeta. Asimismo, es indispensable que las entidades financieras den al evento la importancia que en realidad tiene. Después de Mar del Plata el panorama botánico de América Latina deberá ser mucho más brillante porque podrá ser enfrentado en conjunto y con metas precisas.
La Convención sobre la biodiversidad también tiene entre sus objetivos, según el artículo primero, lo siguiente:
“…el acceso apropiado a los recursos genéticos y a la transferencia de tecnologías apropiadas, teniendo en cuenta todos los derechos sobre esos recursos y esas tecnologías, y el financiamiento apropiado”.
Esperamos que este objetivo se cumpla al pie de la letra.
|
|
|
|
||||||||
|
Nota
Basado en la Conferencia Magistral presentada en el XII Congreso Mexicano de Botánica, Mérida, Yucatán, 5 de octubre de 1993.
|
|||||||||||
|
Referencias Bibliográficas
Campbell, D. G., 1989, “The Importance of Floristic Inventory in the Tropics”, in Campbell, D. G. & D. Hammond (eds.) Floristic Inventory of Tropical Countries, New York Botanical Harden, Bronx, N.Y., pp. 6-30.
Henderson, A., S. P. Churchill & J. L. Luteyn, 1991, “Neotropical plant diversity: Are the northern Andes richer than the Amazon Basin?” Nature 351:21-22. Prance, G. T., 1977, “Floristic inventory in the tropics: where do we stand?” Ann. Missouri Bot. Gard. 64: 659-684. Prance, G. T. & D. G. Campbell, 1988, “The present state of tropical floristics”, Taxon 37(3):519-548. Robinson, N. A., 1993, After Earth in Rebellion. Summit: Where do we go from here? (Environmental Lecture Series). The New York Botanical Garden, Bronx, N.Y. Toledo, V. M. & V. Sosa, 1993, “Floristics in Latin America and the Caribbean: an evaluation of the numbers of plant evaluations and botanists”, Taxon 42:355-364. |
|||||||||||
| ______________________________ | |||||||||||
|
Enrique Forero
Director del Institute of Systematic Botany,
The New York Botanical Garden, Bronx,
Nueva York 10458-5126, Estados Unidos.
|
|||||||||||
| ____________________________________ |
|||||||||||
|
cómo citar este artículo →
Forero, Enrique. 1994. El futuro de la botánica en América Latina. Acuerdos y realidades. Ciencias, núm. 34, abril-junio, pp. 35-41. [En línea].
|
|||||||||||
 |
|
|||||||||||
| José Seade Kuri |
||||||||||||
|
“El porvenir es tan irrevocable como el rígido ayer.
No hay una cosa que no sea una letra silenciosa
de la eterna escritura indescifrable
cuyo libro es el tiempo…”
Jorge Luis Borges, El libro de las Mutaciones.
|
|
|||||||||||
|
En palabras más técnicas y menos poéticas, lo que estos
versos nos dicen es que el mundo en que vivimos es en sí mismo un sistema dinámico. La dificultad para comprenderlo estriba en nuestro reducido entendimiento de las leyes que lo rigen.
Los sistemas dinámicos son un área “joven” de las matemáticas, aunque se remontan a Newton con sus estudios sobre mecánica celeste, y a H. Poincaré, quien inició el estudio cualitativo de las ecuaciones diferenciales. Sin embargo, fue hace apenas unos 30 años que los sistemas dinámicos se establecieron como un área propiamente dicha, gracias al trabajo de matemáticos destacados como S. Smale, Sinai, Lyapunov, A. Douady, M. Herman, D. Sullivan, V. I. Arnold y muchos más.
Actualmente hay una “explosión” de esta área de estudio a nivel mundial, en muchos contextos diferentes. Una característica fascinante de los sistemas dinámicos es la profunda interacción que tienen con otras áreas de las matemáticas y del conocimiento, como la física, la química, la biología y la economía. De hecho, los sistemas dinámicos juegan ya un papel importante inclusive en el arte, a través de los maravillosos conjuntos fractales que aparecen en el estudio de la dinámica de ciertas funciones del plano complejo: los conocidos conjuntos de Julia y de Mandelbrot.
Tratando de precisar el concepto de sistemas dinámicos, podemos decir burdamente que es el estudio de fenómenos deterministas, es decir, consideramos situaciones que dependen de algún parámetro dado, que frecuentemente suponemos es el tiempo, y que varían de acuerdo con leyes establecidas. De manera que el conocimiento de la situación en un momento dado, nos permite reconstruir el pasado y predecir el futuro.
Si queremos ser formales, podemos decir que un sistema dinámico es una familia infinita de funciones (homeomorfismos locales) de un espacio (métrico) en sí mismo, cerrada bajo composiciones, siempre que éstas tengan sentido.
Daré algunos ejemplos que espero ayuden a precisar la definición de sistemas dinámicos y la manera en que estos nos ayudan a entender el mundo que nos rodea.
EJEMPLO 1. Supongamos que, extrañamente, nos encontramos en una “crisis económica” y algún banco ofrece prestarnos dinero. La tasa de interés que cobra el banco es de 2% mensual, y nuestra capacidad real de pago es de dos mil nuevos pesos mensuales máximo. ¿Cuánto dinero queremos que nos preste el banco? Ingenuamente podríamos responder “pues todo lo que se pueda”, pero vamos a analizar este sencillo ejemplo un poco más detenidamente. Denotemos por A0 la cantidad de dinero que le querernos pedir prestado al banco, y por An nuestra deuda después de n meses. Entonces tenemos:
A1 = A0 + 0.02 A0 − 2000
= (1.02) A0 − 2000, A2 = 1.02 ( ) A1 − 2000 = (1.02)2 A0 − 2000(1.02) − 2000, . . . . An = (1.02)n A0 − 2000 (1.02n −1 +…+1.02 +1). Observemos que si A0, el préstamo inicial es de $100 mil nuevos pesos entonces
A1 = (1.02) ⋅ 100,000 − 2000 = A0 ,
A2 = (1.02) ⋅ 100,000 − 2000 = A0 , etcétera, es decir que A0 = $100,000.00 es un punto fijo, o punto de equilibrio, del sistema dinámico en cuestión. Podemos entonces concluir:
a) Si el banco nos presta menos de $100,000.00, algún día terminaremos de pagarle.
b) Si el banco nos presta más de $100,000.00, algún día tendremos que venderle el alma al diablo, o hacer algo para poder pagar.
c) Si el banco nos presta exactamente $100,000.00, simplemente le pagaremos 2 mil pesos mensuales de interés por el resto de nuestra existencia.
Así que conocer las leyes que rigen al sistema, nos permite predecir el futuro. Ustedes tienen la palabra: ¿cuánto quieren que les preste el banco?
Expresiones del tipo presentado, donde tenemos una sucesión de valores {An}, n ∈ Ζ, tales que el valor An está determinado por los valores anteriores An-1, An-2, etcétera, se llaman ecuaciones en diferencias, y dan ejemplos de sistemas dinámicos discretos, donde la palabra discreto significa que el parámetro “tiempo” lo consideramos así: cada mes, cada año, cada hora, etcétera. Las ecuaciones en diferencias, o sistemas dinámicos discretos, más “sencillos” y tal vez más importantes, surgen mediante la iteración de funciones. Regresaremos a este tema al final del artículo.
EJEMPLO 2. Se sabe que ciertas especies animales se multiplican, en condiciones ideales (comida en abundancia, sin luchas internas, etcétera), de manera tal que el crecimiento de la población es proporcional a la cantidad de miembros de la especie (1). Es decir que si P(t) denota la población en el tiempo t, y P’(t) es la velocidad con la que varía la población, entonces existe una constante a > 0 tal que
(I) P’(t) = aP(t),
para todo t > 0, donde 0 denota el tiempo a partir del cual comenzarnos a hacer nuestra observación, y la constante a depende de la población en cuestión y del medio ambiente. Esta ecuación es el ejemplo más sencillo e importante de lo que es una ecuación diferencial. Es fácil ver que cualquier función que satisface la ecuación diferencial (I) necesariamente es de la forma
(II) Y(t) = Keat
donde K es una constante, determinada por “la condición inicial”. En nuestro caso K es la población inicial en t = 0. La ley de crecimiento de población dada por la ecuación (I) se conoce como Ley Malthusiana de crecimiento, y la ecuación (II) nos dice que si la especie en cuestión se rige por esta ley de crecimiento, entonces, independientemente de la población inicial, si P(0) > 0, la población tenderá a infinito exponencialmente. La experiencia nos enseña que esto no sucede en la realidad por tiempo prolongado, así que debe haber otros factores a considerar. Una vez que la población sobrepasa un cierto número, comienza a haber escasez de alimentos, se producen luchas internas, desechos contaminantes, etcétera. Esto nos lleva a considerar la “Ley logística” de crecimiento de poblaciones:
P’(t) = aP(t) - bP2(t)
donde a, b son constantes positivas y a es mucho mayor que b. Las constantes a y b son los coeficientes vitales de la especie. Obsérvese que si la población P(t) es “pequeña”, el término bP2(t) es despreciable, pues b es muy pequeña en relación con a, por lo que el crecimiento asemeja al regido por la ley malthusiana. Sin embargo, al aumentar P(t), la contribución bP2 crece en forma cuadrática, por lo que juega un papel importante.
Esta ecuación diferencial es de primer orden, del tipo “separable” y por tanto fácil de resolver; si P(0) es la población inicial en t = 0, entonces (por el teorema de existencia de unicidad de soluciones con condiciones iniciales) existe una única función de t que satisface la ecuación diferencial y toma el valor P(0) en t = 0. La solución buscada es:
Si hacernos L = a/b, observamos que P(t) tiende a L cuando t tiende a ∞, independientemente de la población inicial, pues e-at tiende a 0 cuando t tiende a infinito. Además, P(t) es una función monótona creciente para t > 0. Más aún, dado que
P’’(t) = aP’(t) - 2bP(t) · P’(t),
se ve que P’ es creciente para P(t) < L/2 y es decreciente para P(t) > L/2, con un punto de inflexión en P(t) = L/2. Por tanto, la gráfica de P es del tipo: (ver figura 1).
A partir de esta información concluimos que el periodo de tiempo antes de que la población alcance la mitad de su límite L, es un periodo de crecimiento acelerado, semejante al crecimiento regido por la ley malthusiana. Después de este punto, la tasa de crecimiento disminuye hasta llegar a cero.
De esta forma la ley logística nos permite hacer predicciones sobre el crecimiento de las especies, predicciones que se han comprobado en diversos estudios y experimentos. Por ejemplo, en 1920 los científicos norteamericanos Pearl y Reed publicaron un artículo con predicciones sobre el crecimiento de la población (humana) en los Estados Unidos de Norteamérica. Para ello estudiaron el crecimiento de dicha población desde 1790, y así estimaron los coeficientes vitales a y b como: a = 0.03134 y b = (1.5887)10-10. Considerando que la población de los Estados Unidos en 1920 era de 105711000 habitantes, la ley logística de crecimiento indicaba que para 1950, la población sería de 149053000, y la población real era de 150697000, lo cual representa un error de 1.1%, que ¡no está mal!
Obsérvese que la ley logística
P’(t) = aP(t) - b2P(t),
puede escribirse en la forma
donde L = a/b es el límite de la población. El término aP(t) es el potencial biótico de la especie, es decir, la tasa potencial de crecimiento en condiciones ideales, y el término (L - P)/L es la resistencia ambiental al crecimiento.
Ahora bien, supongamos que tenernos dos especies P1 y P2, cuyo crecimiento se rige por la ley logística cuando las especies están aisladas de otras. ¿Qué sucede si ahora juntamos a las especies P1 y P2, de manera tal que tienen que competir entre sí por el alimento, espacio, etcétera? En este caso las ecuaciones se transforman en:
donde α1 y α2 son constantes que indican el grado de influencia de una especie en la otra. Nos interesan preguntas como las siguientes:
1. ¿Existen constantes c1, c2 tales que P1(t) ≡ c1 y P2(t) ≡ c2 satisfacen el sistema de ecuaciones anterior? Si tales valores existen, se les llama puntos de equilibrio del sistema. Obsérvese que y constantes equivalen a pedir que P1 y P2 sea 0, y significa que las dos especies puedan coexistir permanentemente.
2. Supongamos que (c1, c2) es un punto de equilibrio del sistema, y súbitamente agregamos algunos miembros de la primera especie, ¿permanecerán P1 y P2 cerca del valor (c1, c2) para todo tiempo futuro? Puede suceder, por ejemplo, que los miembros adicionales de la primera especie le den a ésta una ventaja tal sobre la segunda especie, de manera que ésta tiende a la extinción mientras que P1 tiende a un valor límite L1. Si esto sucede, decimos que el punto (c1, c2) es inestable, mientras que si P1 y P2 permanecen cerca de (c1, c2) para todo tiempo futuro, decirnos que éste es un punto de equilibrio estable. Más aún, puede suceder que si agregamos algunos miembros de cualquiera de las dos especies, al pasar el tiempo las poblaciones P1 y P2 tienden otra vez a la solución de equilibrio (c1, c2). En este caso se dice que (c1, c2) es un atractor.
3. Supongamos que P1 y P2 tienen valores arbitrarios para t igual a 0. ¿Qué ocurre cuando el tiempo tiende a infinito? ¿Triunfará alguna de las dos especies, tenderán a una solución de equilibrio?
Si en vez de dos especies en competencia, consideramos tres especies, o n especies en competencia, lo que obtendremos es un sistema de ecuaciones de la forma:
 donde f1,…, fn son funciones de P1(t),…, Pn(t), t. Esto es lo que se conoce como un sistema de n ecuaciones diferenciales de primer orden en P1,…, Pn. A un conjunto de funciones que satisfacen este sistema se le llama una solución. Parte fundamental de estos sistemas dinámicos es el estudio de las propiedades cualitativas de las soluciones, donde por propiedades cualitativas entendernos propiedades del tipo de las mencionadas en las preguntas 1-3.
Dado un sistema de ecuaciones diferenciales
 una solución del sistema es una n-ada (ø1,…,øn) de funciones de t que satisfacen el sistema. Es decir que ø = (ø1,…,øn) satisface:
 Por ejemplo, la pareja
(e2t + e -3t, 2e2t - 3e-3t),
es una solución del sistema lineal,
x'1 = x2
x'2 = 6x1 - x2 ya que,
y,
Obsérvese que para cada t, la n-ada (ø1(t),…,øn(t)) es un punto en Rn. Así, al variar el tiempo, la solución (ø1,…,øn) describe una curva, o trayectoria, en Rn. En este caso a Rn se le llama el espacio fase, y la curva que describe la solución en Rn se llama una órbita del sistema. Las soluciones de equilibrio corresponden al caso donde las derivadas x’1,…,x’n son cero para todo t, así que la órbita consiste de solamente el punto en el equilibrio. El teorema fundamental de ecuaciones diferenciales es el teorema de existencia y unicidad de soluciones, el cual nos dice que si las funciones f1,…,fn son funciones diferenciables de clase c1, entonces:
a) Las órbitas son curvas simples, es decir, sin auto-intersecciones.
b) Las órbitas son ajenas dos a dos: si dos órbitas se intersectan en un punto entonces son idénticas. Por cada punto del espacio fase pasa una y solo una órbita del sistema.
c) las órbitas llenan todo el espacio Rn.
Luego, dado un sistema de ecuaciones como el anterior, sus órbitas descomponen al espacio fase en unión ajena de curvas simples y puntos de equilibrio. A esta descomposición de Rn se le llama el retrato fase del sistema dinámico. Nos preguntamos cuestiones como: ¿Existen puntos de equilibrio? ¿Cuántos hay? ¿Existen órbitas periódicas? ¿Cuántas?
(Estas son soluciones del sistema tales que existen t0 y T > 0, satisfaciendo Φ(t0) = Φ(t0 + T), donde Φ = (Φ1,…,Φn)) ¿Son estables las soluciones?, es decir que si Φ(t) y ψ(t) son soluciones tales que para un cierto tiempo t0 los puntos Φ(t0) y ψ(t0) están “muy cercanos”, entonces Φ(t) y ψ(t) están “muy cercanos” para todo t.
¿Cuáles son los conjuntos límite de estas órbitas?, o en otras palabras ¿dónde se acumulan o dónde nacen y dónde mueren las órbitas?, etcétera.
El estudio de éstas y otras preguntas relacionadas, constituye el tema central de los sistemas dinámicos.
EJEMPLO 3. Considérese el sistema de ecuaciones en R2 definido por:
 con k constante. Cualquier solución de este sistema es de la forma
donde c1 y c2 son constantes arbitrarias. Hay 5 casos cualitativamente distintos, dependiendo de k:
caso 1: k >1.
caso 2: k=1. caso 3: 1>k>0. caso 4: k=0. caso 5: k<0. En todos los casos, el origen 0 ∈ R2 es punto de equilibrio del sistema, si k ≠ 0, este es el único punto de equilibrio, pero si k = 0, todos los puntos en el eje “y” son puntos de equilibrio. En los otros casos, hay 5 órbitas especiales: el punto de equilibrio 0, y los 4 semiejes (x > 0, y = 0), (x < 0, y = 0), (x = 0, y > 0) y (x = 0, y < 0). Los retratos fase son en cada caso, como se indica en la figura 2.
En todos los casos, las órbitas que no son puntos fijos tienden a infinito. En los casos, 1, 2, 3 y 5, todas las órbitas nacen en el origen.
También podemos considerar sistemas dinámicos que dependen de un parámetro discreto, como es el caso de las ecuaciones en diferencias, mencionadas anteriormente. Aquí también podemos hablar de puntos de equilibrio, órbitas periódicas, atractores, repulsores, etcétera. Si en una ecuación en diferencias, hacemos que nuestro intervalo de tiempo sea menor, por ejemplo en lugar de “cada mes” consideramos cada semana, o cada día, o cada hora, etcétera, nuestra ecuación en diferencias se va transformando hasta que, cuando la longitud de los intervalos, tiende a cero, lo que nos queda en el límite, si éste existe, es una ecuación diferencial.
EJEMPLO 4. Supongamos que ya superamos nuestra “crisis económica” del ejemplo 1, y queremos abrir una cuenta de inversión en el banco. Nuestra inversión inicial es de C0 pesos, y el banco paga, por decir algo, el 10% de interés anual, que se reinvierte en nuestra cuenta anualmente. Nos preguntamos cuánto tendremos después de n años. Para responder a esta pregunta, denotamos por C(n) nuestro capital después de n años, así que C(0) = C0. Después de un año tendremos C(1) = C0 + (0.1) C0 = (1.1)C0. Al segundo año tendremos C(2) = C(1) + (0.1)C(1) = (1.1) C(1) = (1.1)2C0 y así sucesivamente. Luego, C(n) = (1.1)nC0, lo que responde nuestra pregunta original. Más generalmente, si el interés que nos paga el banco es 100 · I%, lo que tendremos después de n años es (1 + I)nC0. Nos preguntamos ahora qué sucede si cambiamos el tipo de inversión que tenemos. Denotemos por F0 nuestro capital inicial, el banco nos paga 100 · 1% de interés anual, pero la diferencia estriba en que el interés que ganamos se reinvierte en la cuenta m veces al año. Sea h = 1/m la fracción del año que consiste de un periodo de reinversión del interés. (Por ejemplo h = 1/12 si el interés se reinvierte mensualmente). Denotemos por n el número de periodos de reinversión considerado. (Así, si h es 1/12 y n es 30, el periodo de tiempo considerado es de dos años y medio).
Entonces tenemos:
F(1) = F0 + ( I / m)F0 = (1 + I / m) F0,
F(2) = F(1) + ( I / m) F(1) = (1 + I / m) F(1) = (1 + I / m)2 F0, .
. . F(n) = F(n-1) + ( I / m) F(n-1) = ...= (1 + I / m)n F0. o equivalentemente
F(n+1) - F(n) = I / m F(n).
Es decir, recordando que h denota la fracción del año que consiste de un periodo de reinversión del interés,
F(n + h) - F(n) = IhF (n),
y dividiendo ambos lados de la ecuación por h = 1/m, obtenemos,
haciendo m, el número de periodos de reinversión, tender a infinito nos queda
es decir, escribiendo t en vez de n, obtenemos
F’(t) = I · F(t)
que es una ecuación diferencial del tipo f’(t) = af(t), considerado en la ley de crecimiento de Malthus, del ejemplo 2.
Un tipo importante de ecuaciones en diferencias son las llamadas ecuaciones autónomas de primer orden. Estas son las ecuaciones en diferencias de la forma
An + 1 = f(An),
donde f es una función de un conjunto Ω en sí mismo. Así que si hacemos,
f1 = f,
f2 = f o f, . . . fn =( f)( f) ...(f), n veces, entonces An es el n-ésimo iterado de A0,
An = fn(A0).
Es decir que dada una función f de Ω en sí mismo, nos podemos fijar en todas sus iteraciones {fn}, y en el sistema dinámico que éstas nos definen. Fascinantes ejemplos y aplicaciones surgen al considerar este tipo de sistemas dinámicos. En cierto sentido, estudiar la dinámica de difeomorfismos en espacios fase de dimensión n es “equivalente” a estudiar la dinámica de ecuaciones diferenciales en espacios fase de dimensión n + 1. Por un lado, dadas las órbitas de una ecuación diferencial en, digamos Rn + 1, nos podemos fijar en pequeñas “transversales” a las órbitas, que son discos de dimensión n, y la ecuación diferencial determina difeomorfismos locales entre estas transversales, los cuales definen el llamado “Mapeo de Poincaré” y el “grupo de holonomía” de la ecuación. Recíprocamente, cualquier difeomorfismo F de un espacio fase X en dimensión n, determina un flujo, o ecuación diferencial, en un espacio fase Y de dimensión n + 1, a través de una construcción sencilla llamada “la suspensión”, y el mapeo de Poincaré de la ecuación diferencial en Y es precisamente el difeomorfismo F. Estas dos construcciones están bien estudiadas y sugerimos al lector interesado consultar la bibliografía (2).
Otro ejemplo muy interesante es el de la “familia cuadrática”, fc: C → C definida para cada constante c ∈ C, por f(z) = z2 + c. Para cualquier c fija se tiene que ∞ es un punto fijo atractor, pues |z2| = |z|2, así que la norma de f(z) crece cuadráticamente. Esto significa que si |z| es suficientemente grande (con respecto a c), entonces
Con puntos de norma pequeña, puede suceder que el punto tienda a infinito o no. Puede entonces suceder que el 0 ∈ C tienda a ∞ o no, dependiendo del parámetro c. El Conjunto de Mandelbrot (figura 3) es el conjunto de valores del parámetro c ∈ C para los cuales la sucesión {fck (0)} no tiende a infinito. Más sobre este ejemplo puede verse en (3), que tiene un tratamiento muy elemental y accesible. Para una exposición general de la dinámica de la familia cuadrática, véase (4).
|
|
|
||||||||||
|
Referencias Bibliográficas
1. M. Braun. 1991, Ecuaciones diferenciales y sus aplicaciones, Versión en español publicada por Editorial Interamericana.
2. J. Palis & W. de Melo, 1982, Geometric theory of dynamical systems, Springer Verlag, Graduate Texts in Mathematics. 3. J. T. Sandefur, 1990, Discrete Dynamical Systems, theory and applications, Oxford University Press. 4. Blanchard, 1984, Complex Analytic Dynamics on the Riemann Sphere, Bull. A. M. S. 11, No. 1 p.85-141. 5. M. Hirsch y S. Smale, 1974, Differential equations, dynamical systems and linear algebra, Academic Press. 6. H. O. Peitgen, 1986, The beauty of fractals, Springer Verlarg. |
||||||||||||
| __________________________________ | ||||||||||||
|
José Seade Kuri
Instituto de Matemáticas, Universidad Nacional Autónoma de México
y Instituto Tecnológico Autónomo de México.
|
||||||||||||
| __________________________________ | ||||||||||||
|
cómo citar este artículo →
Seade Kuri, José. 1994. Una introducción a los sistemas dinámicos. Ciencias, núm. 34, abril-junio, pp. 23-29. [En línea].
|
||||||||||||


